

First human model of in vitro Candida albicans persistence within granuloma for the reliable study of host-fungi interactions
- PMID: 22768252
- PMCID: PMC3387014
- DOI: 10.1371/journal.pone.0040185
First human model of in vitro Candida albicans persistence within granuloma for the reliable study of host-fungi interactions
Abstract
Background: The balance between human innate immune system and Candida albicans virulence signaling mechanisms ultimately dictates the outcome of fungal invasiveness and its pathology. To better understand the pathophysiology and to identify fungal virulence-associated factors in the context of persistence in humans, complex models are indispensable. Although fungal virulence factors have been extensively studied in vitro and in vivo using different immune cell subsets and cell lines, it is unclear how C. albicans survives inside complex tissue granulomas.
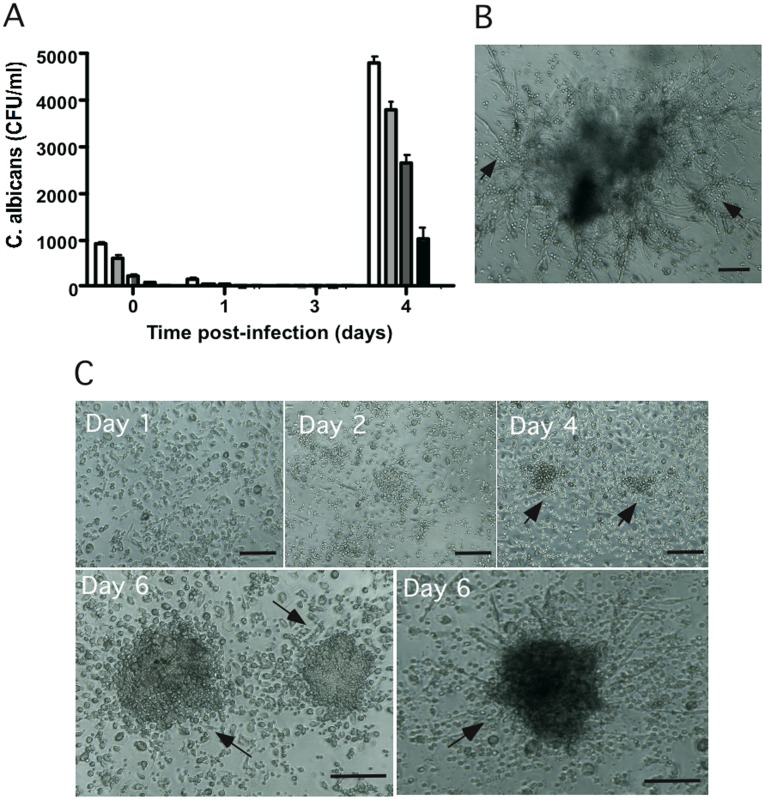
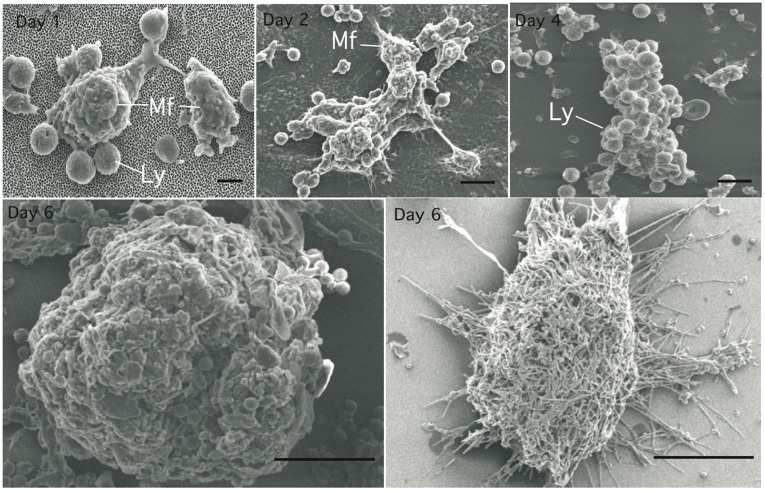
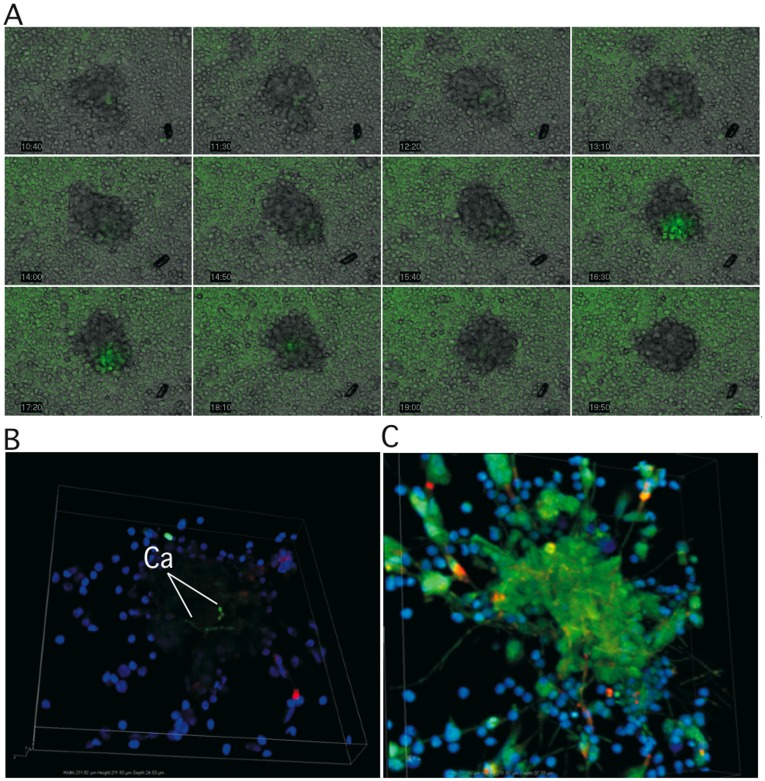
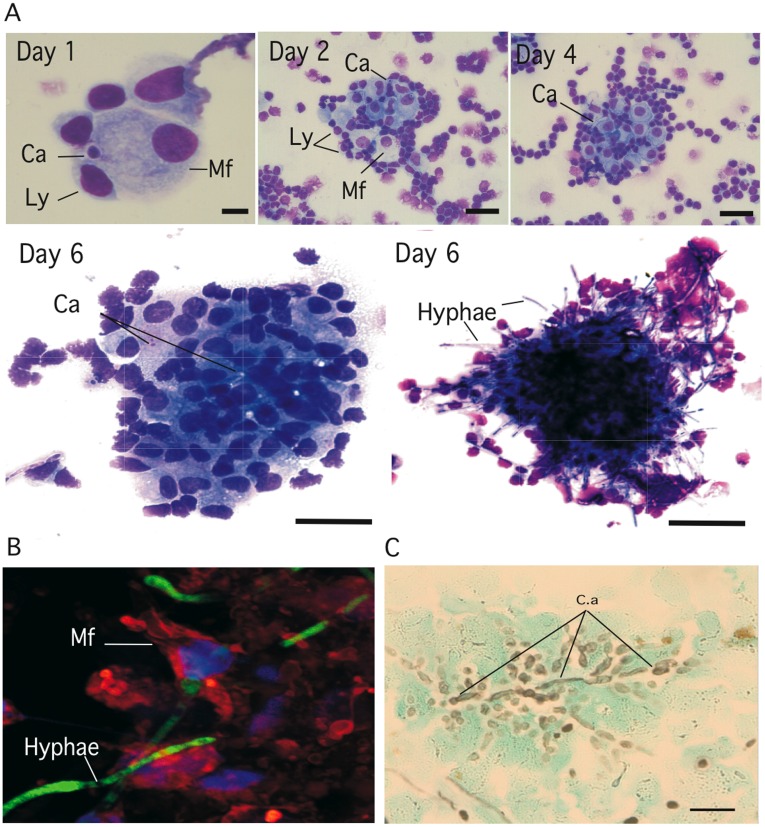
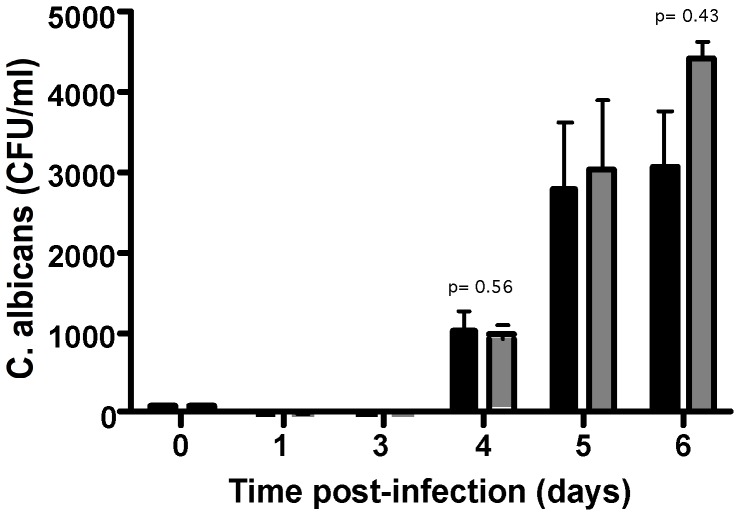
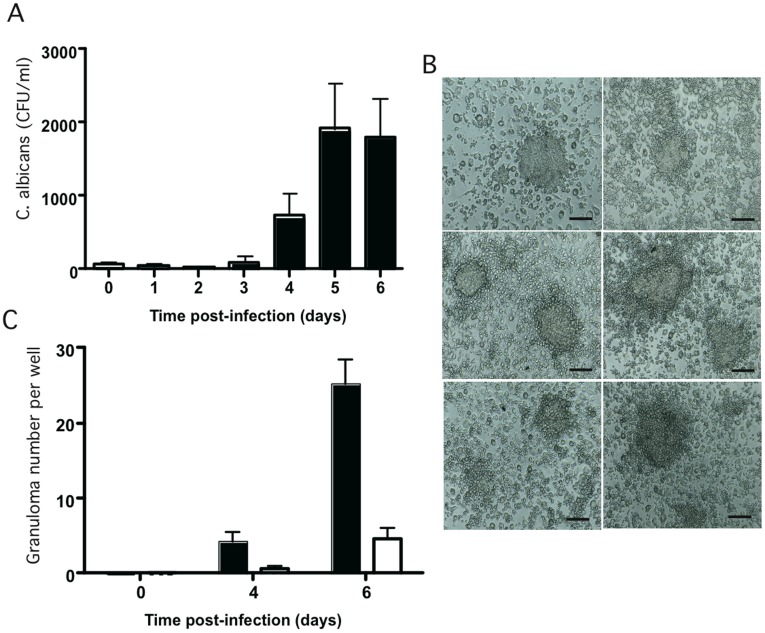
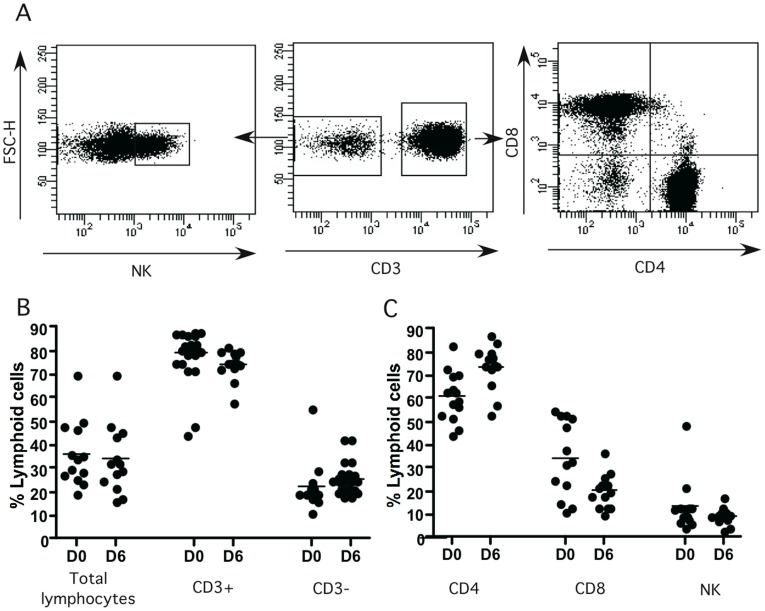
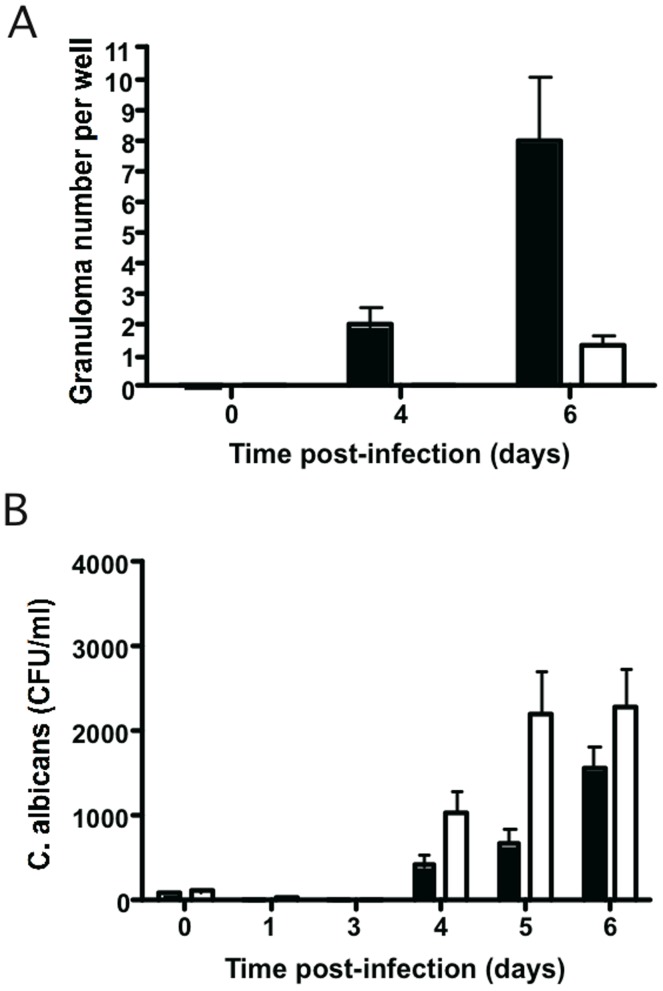
Methodology/principal finding: We developed an original model of in vitro human granuloma, reproducing the natural granulomatous response to C. albicans. Persistent granulomas were obtained when the ratio of phagocytes to fungi was high. This in vitro fungal granuloma mimics natural granulomas, with infected macrophages surrounded by helper and cytotoxic T lymphocytes. A small proportion of granulomas exhibited C. albicans hyphae. Histological and time-lapse analysis showed that C. albicans blastoconidia were located within the granulomas before hyphae formation. Using staining techniques, fungal load calculations, as well as confocal and scanning electron microscopy, we describe the kinetics of fungal granuloma formation. We provide the first direct evidence that C. albicans are not eliminated by immunocompetent cells inside in vitro human granulomas. In fact, after an initial candicidal period, the remaining yeast proliferate and persist under very complex immune responses.
Conclusions/significance: Using an original in vitro model of human fungal granuloma, we herein present the evidence that C. albicans persist and grow into immunocompetent granulomatous structures. These results will guide us towards a better understanding of fungal invasiveness and, henceforth, will also help in the development of better strategies for its control in human physiological conditions.
Conflict of interest statement
Figures
References
-
- Calderone RA, Fonzi WA. Virulence factors of Candida albicans. Trends Microbiol. 2001;9:327–335. - PubMed
-
- Calderone R, Suzuki S, Cannon R, Cho T, Boyd D, et al. Candida albicans: adherence, signaling and virulence. Med Mycol. 2000;38:125–137. - PubMed
-
- Calera JA, Herman D, Calderone R. Identification of YPD1, a gene of Candida albicans which encodes a two-component phosphohistidine intermediate protein. Yeast. 2000;16:1053–1059. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
