

Effects of dimethyl fumarate on neuroprotection and immunomodulation
- PMID: 22769044
- PMCID: PMC3419128
- DOI: 10.1186/1742-2094-9-163
Effects of dimethyl fumarate on neuroprotection and immunomodulation
Abstract
Background: Neuronal degeneration in multiple sclerosis has been linked to oxidative stress. Dimethyl fumarate is a promising novel oral therapeutic option shown to reduce disease activity and progression in patients with relapsing-remitting multiple sclerosis. These effects are presumed to originate from a combination of immunomodulatory and neuroprotective mechanisms. We aimed to clarify whether neuroprotective concentrations of dimethyl fumarate have immunomodulatory effects.
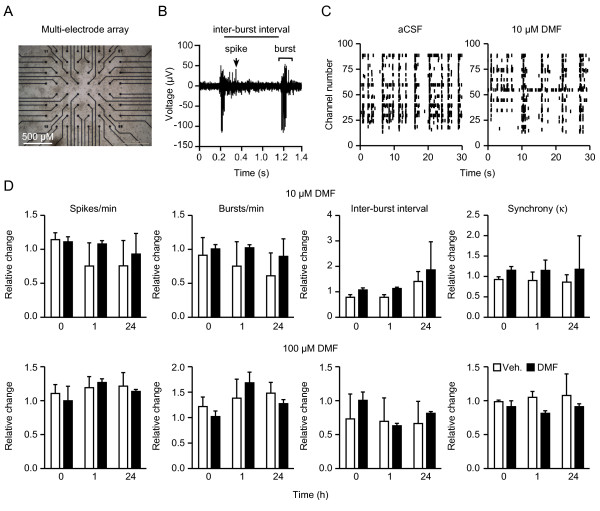
Findings: We determined time- and concentration-dependent effects of dimethyl fumarate and its metabolite monomethyl fumarate on viability in a model of endogenous neuronal oxidative stress and clarified the mechanism of action by quantitating cellular glutathione content and recycling, nuclear translocation of transcription factors, and the expression of antioxidant genes. We compared this with changes in the cytokine profiles released by stimulated splenocytes measured by ELISPOT technology and analyzed the interactions between neuronal and immune cells and neuronal function and viability in cell death assays and multi-electrode arrays. Our observations show that dimethyl fumarate causes short-lived oxidative stress, which leads to increased levels and nuclear localization of the transcription factor nuclear factor erythroid 2-related factor 2 and a subsequent increase in glutathione synthesis and recycling in neuronal cells. Concentrations that were cytoprotective in neuronal cells had no negative effects on viability of splenocytes but suppressed the production of proinflammatory cytokines in cultures from C57BL/6 and SJL mice and had no effects on neuronal activity in multi-electrode arrays.
Conclusions: These results suggest that immunomodulatory concentrations of dimethyl fumarate can reduce oxidative stress without altering neuronal network activity.
Figures




References
-
- Kappos L, Gold R, Miller DH, MacManus DG, Havrdova E, Limmroth V, Polman CH, Schmierer K, Yoursry TA, Yang M, Eraksoy M, Meluzinova E, Rektor I, Dawson KT, Sandrock AW, O’Neill GN. bg-12 Phase IIb Study Investigators. Efficacy and safety of oral fumarate in patients with relapsing-remitting multiple sclerosis: a multicentre, randomised, double-blind, placebo-controlled phase IIb study. Lancet. 2008;372:1463–1472. doi: 10.1016/S0140-6736(08)61619-0. - DOI - PubMed
-
- Gold R, Kappos L, Bar-Or D, Arnold D, Giovannoni G, Selmaj K, Yang M, Dawson K. Clinical efficacy of BG-12, an oral therapy, in relapsing-remitting multiple sclerosis: data from the phase 3 DEFINE trial. , Amsterdam; October 19-22 2011. 17:S9-S52.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
