

Identification of stem cell populations in sweat glands and ducts reveals roles in homeostasis and wound repair
- PMID: 22770217
- PMCID: PMC3423199
- DOI: 10.1016/j.cell.2012.04.045
Identification of stem cell populations in sweat glands and ducts reveals roles in homeostasis and wound repair
Abstract
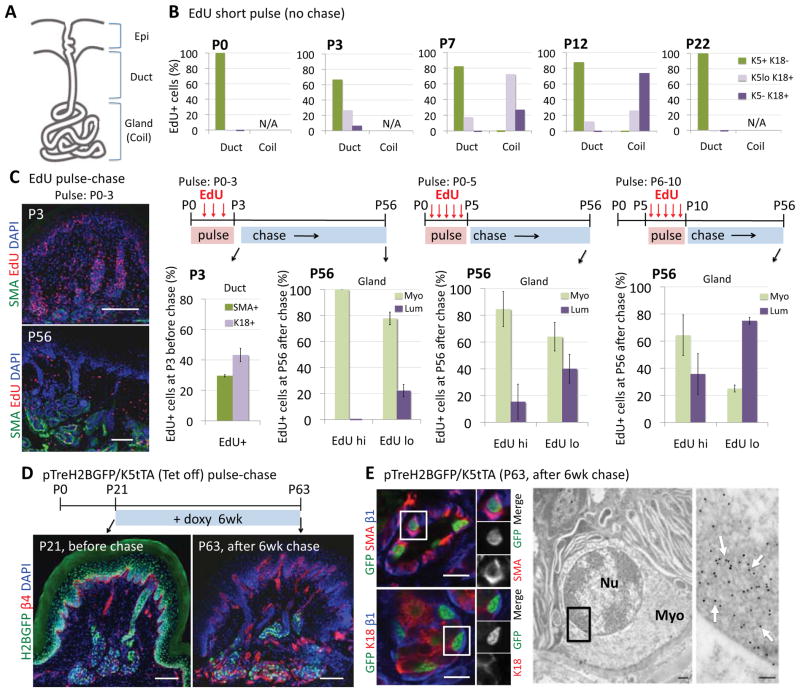
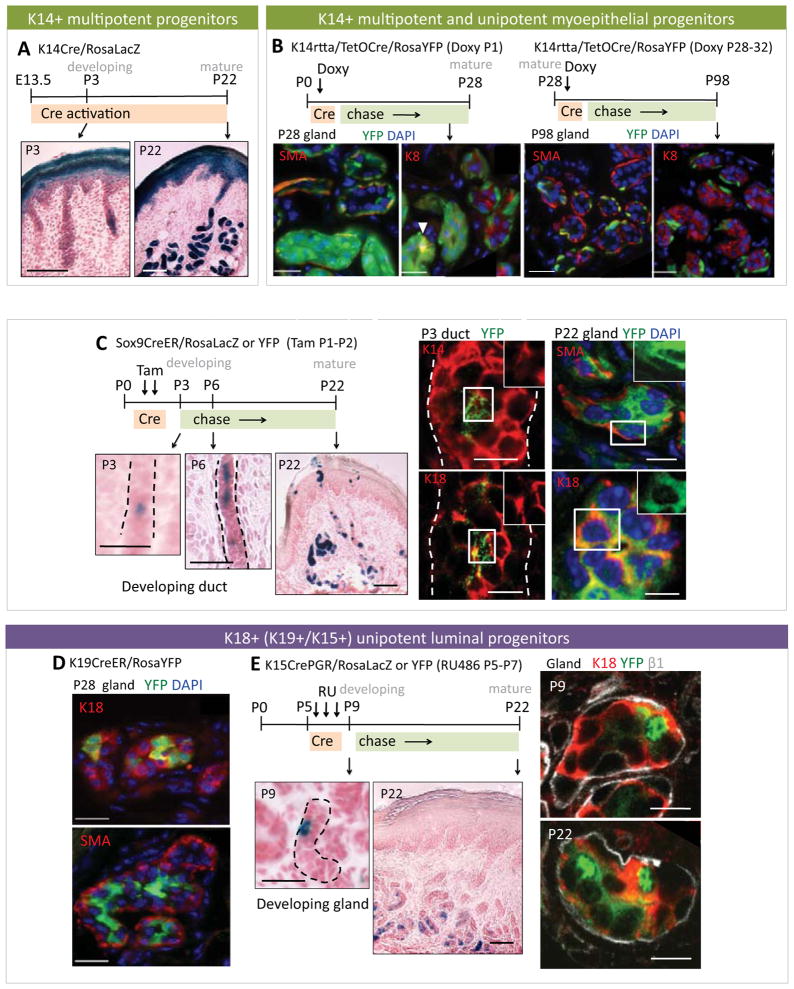
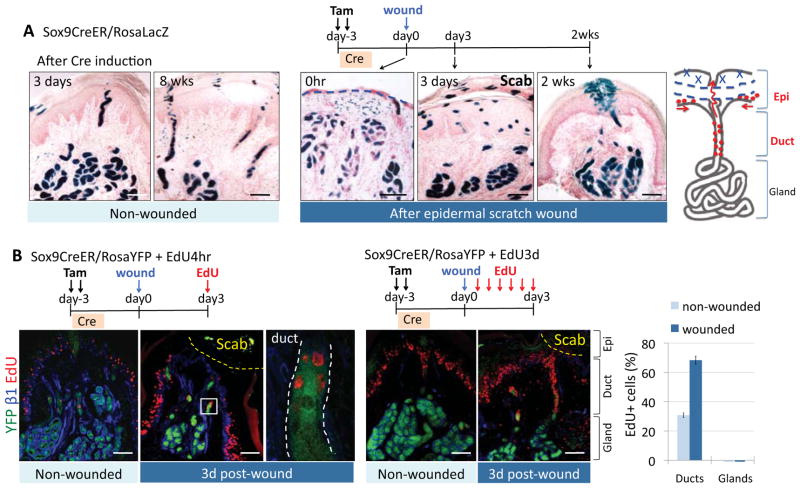
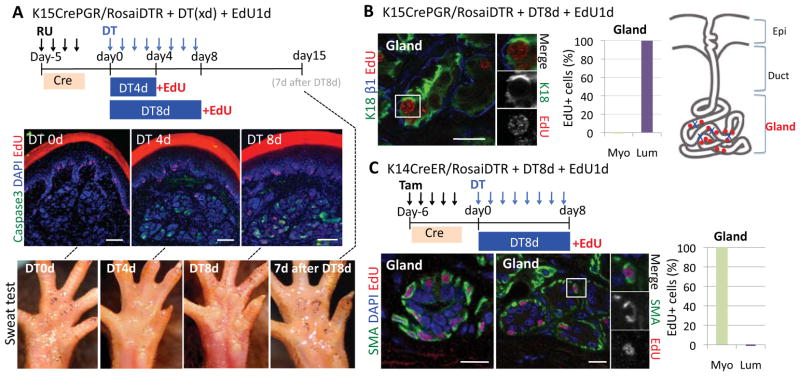
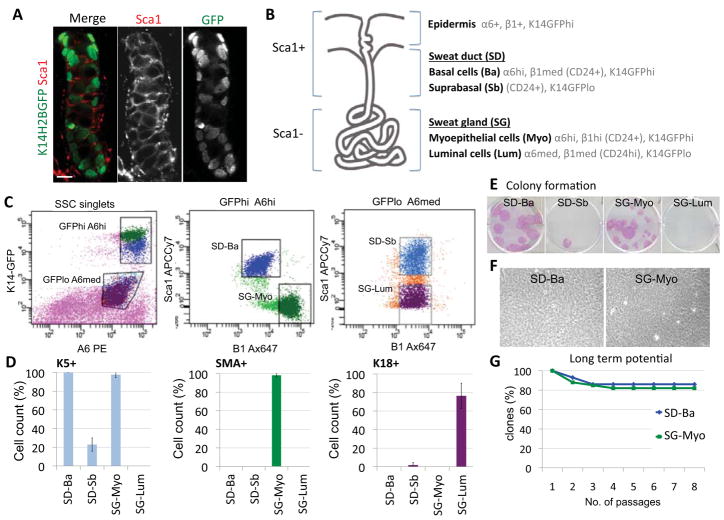
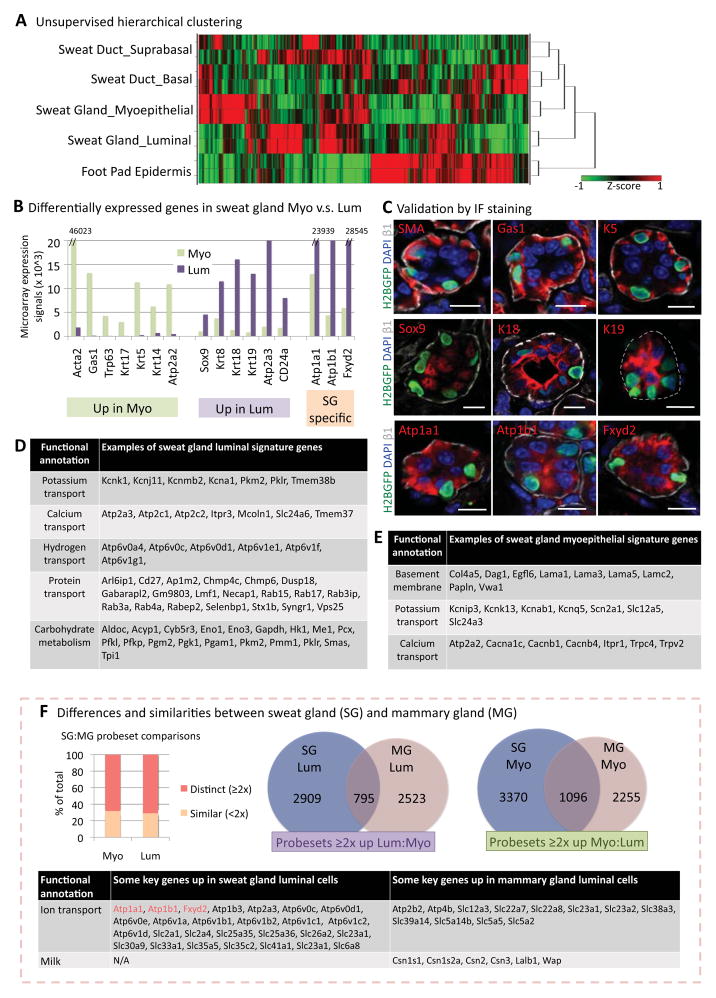
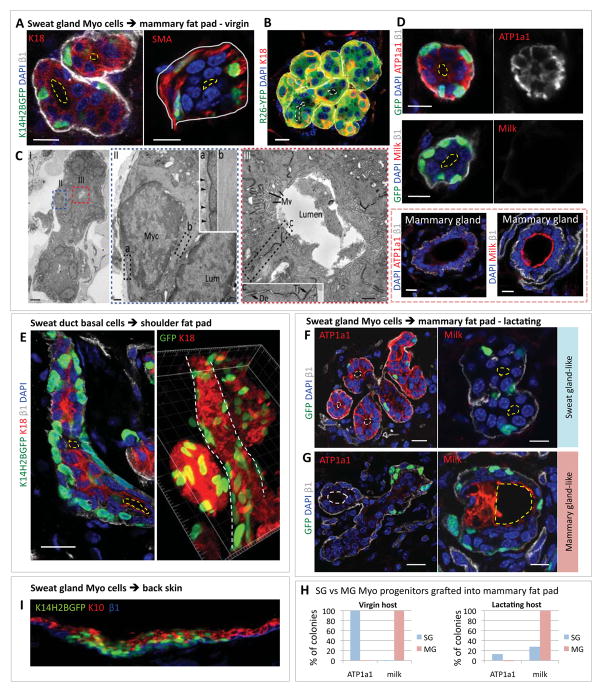
Sweat glands are abundant in the body and essential for thermoregulation. Like mammary glands, they originate from epidermal progenitors. However, they display few signs of cellular turnover, and whether they have stem cells and tissue-regenerative capacity remains largely unexplored. Using lineage tracing, we here identify in sweat ducts multipotent progenitors that transition to unipotency after developing the sweat gland. In characterizing four adult stem cell populations of glandular skin, we show that they display distinct regenerative capabilities and remain unipotent when healing epidermal, myoepithelial-specific, and lumenal-specific injuries. We devise purification schemes and isolate and transcriptionally profile progenitors. Exploiting molecular differences between sweat and mammary glands, we show that only some progenitors regain multipotency to produce de novo ductal and glandular structures, but that these can retain their identity even within certain foreign microenvironments. Our findings provide insight into glandular stem cells and a framework for the further study of sweat gland biology.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Biedermann T, Pontiggia L, Bottcher-Haberzeth S, Tharakan S, Braziulis E, Schiestl C, Meuli M, Reichmann E. Human eccrine sweat gland cells can reconstitute a stratified epidermis. J Invest Dermatol. 2010;130:1996–2009. - PubMed
-
- Blanpain C, Lowry WE, Geoghegan A, Polak L, Fuchs E. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 2004;118:635–648. - PubMed
-
- Blecher SR. Anhidrosis and absence of sweat glands in mice hemizygous for the Tabby gene: supportive evidence for the hypothesis of homology between Tabby and human anhidrotic (hypohidrotic) ectodermal dysplasia (Christ-Siemens-Touraine syndrome) J Invest Dermatol. 1986;87:720–722. - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
