

Adaptive mutations that prevent crosstalk enable the expansion of paralogous signaling protein families
- PMID: 22770222
- PMCID: PMC3415470
- DOI: 10.1016/j.cell.2012.05.033
Adaptive mutations that prevent crosstalk enable the expansion of paralogous signaling protein families
Abstract
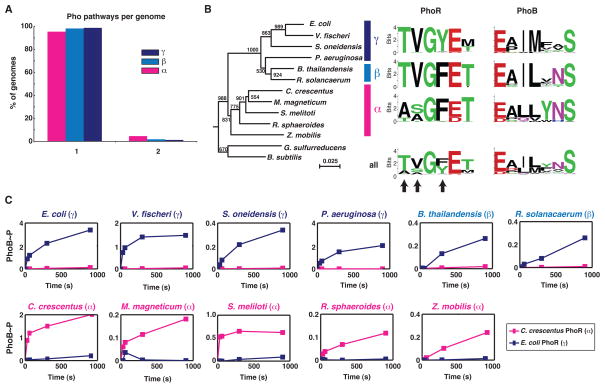
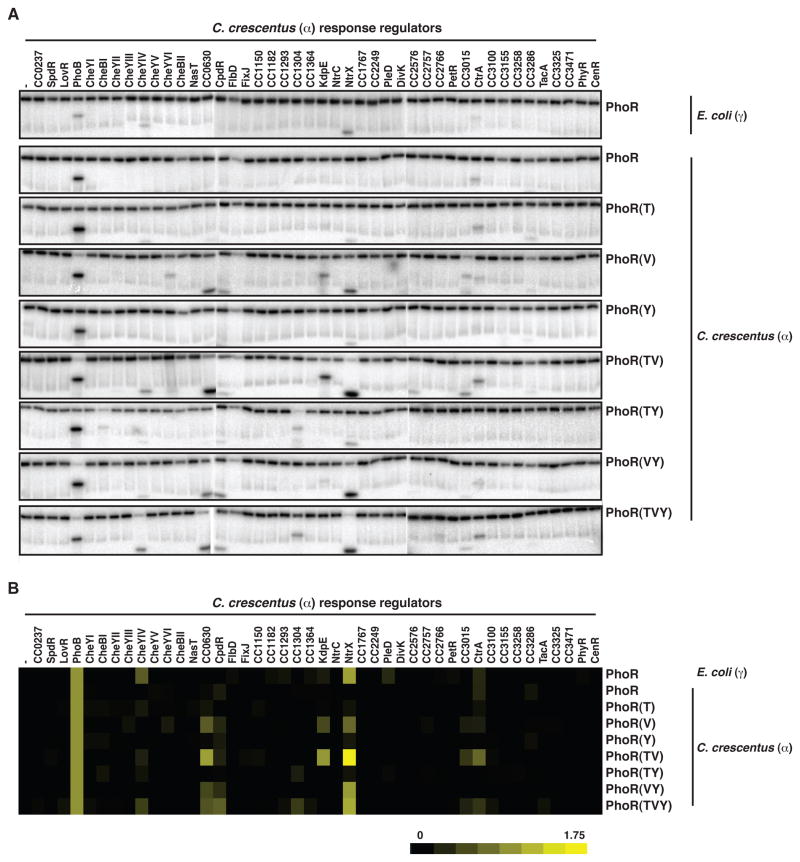
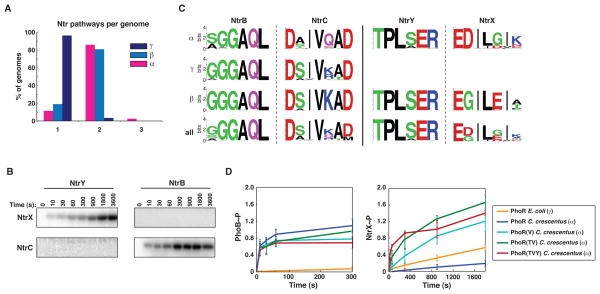
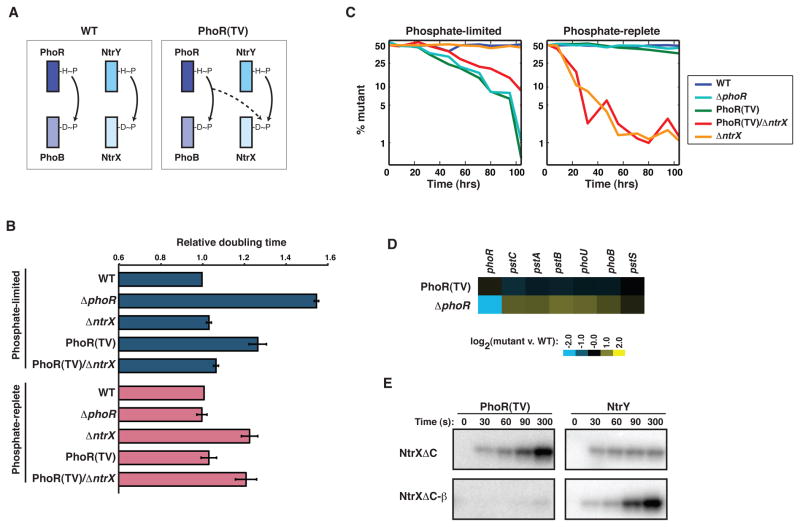
Orthologous proteins often harbor numerous substitutions, but whether these differences result from neutral or adaptive processes is usually unclear. To tackle this challenge, we examined the divergent evolution of a model bacterial signaling pathway comprising the kinase PhoR and its cognate substrate PhoB. We show that the specificity-determining residues of these proteins are typically under purifying selection but have, in α-proteobacteria, undergone a burst of diversification followed by extended stasis. By reversing mutations that accumulated in an α-proteobacterial PhoR, we demonstrate that these substitutions were adaptive, enabling PhoR to avoid crosstalk with a paralogous pathway that arose specifically in α-proteobacteria. Our findings demonstrate that duplication and the subsequent need to avoid crosstalk strongly influence signaling protein evolution. These results provide a concrete example of how system-wide insulation can be achieved postduplication through a surprisingly limited number of mutations. Our work may help explain the apparent ease with which paralogous protein families expanded in all organisms.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
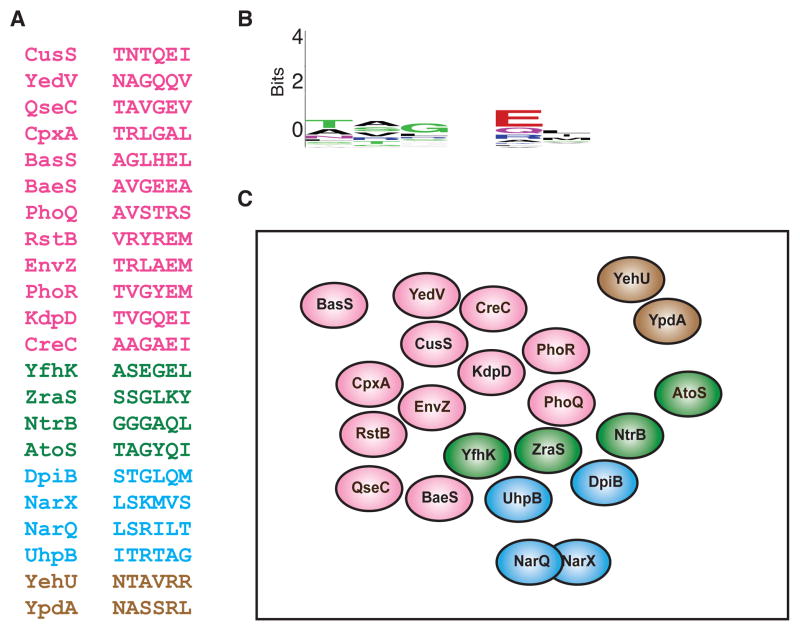
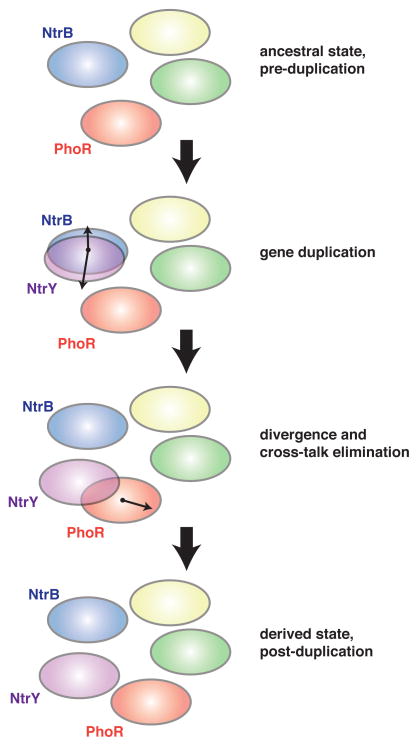
References
-
- Casino P, Rubio V, Marina A. Structural insight into partner specificity and phosphoryl transfer in two-component signal transduction. Cell. 2009;139:325–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
