

Progenitor function in self-renewing human epidermis is maintained by the exosome
- PMID: 22770246
- PMCID: PMC3392610
- DOI: 10.1016/j.stem.2012.04.022
Progenitor function in self-renewing human epidermis is maintained by the exosome
Abstract
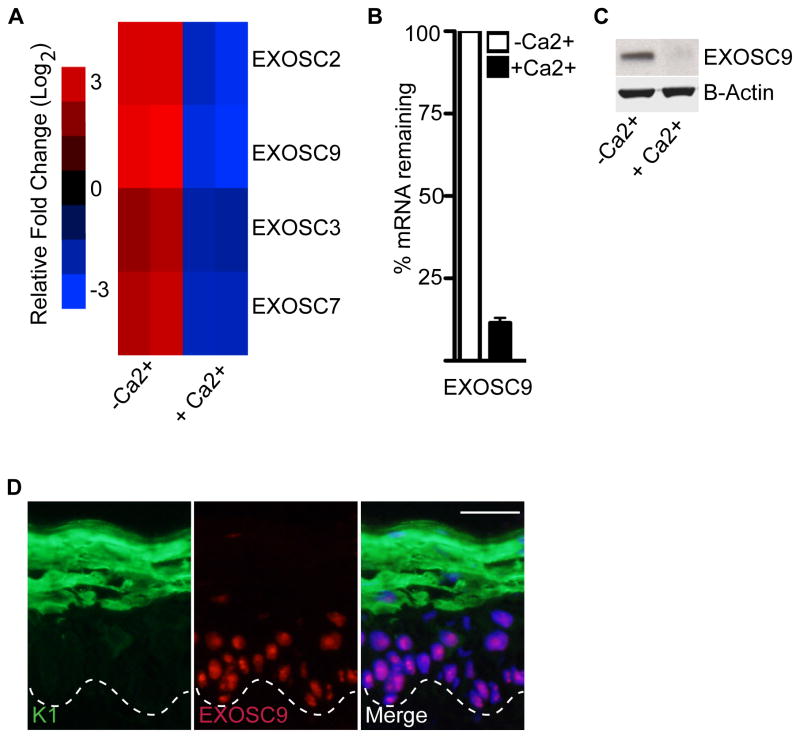
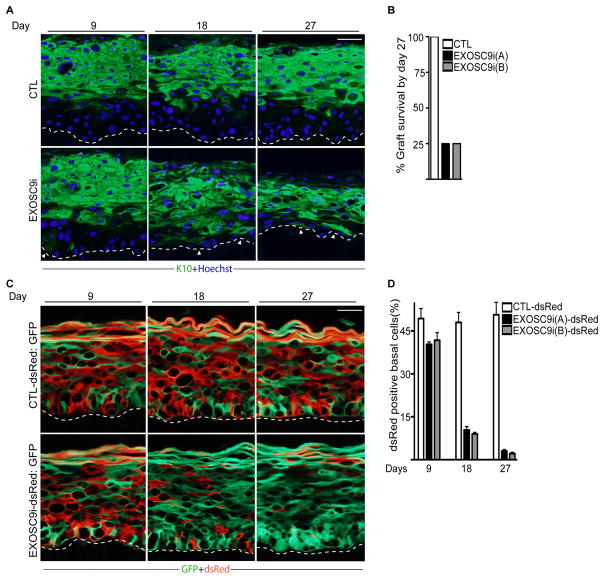
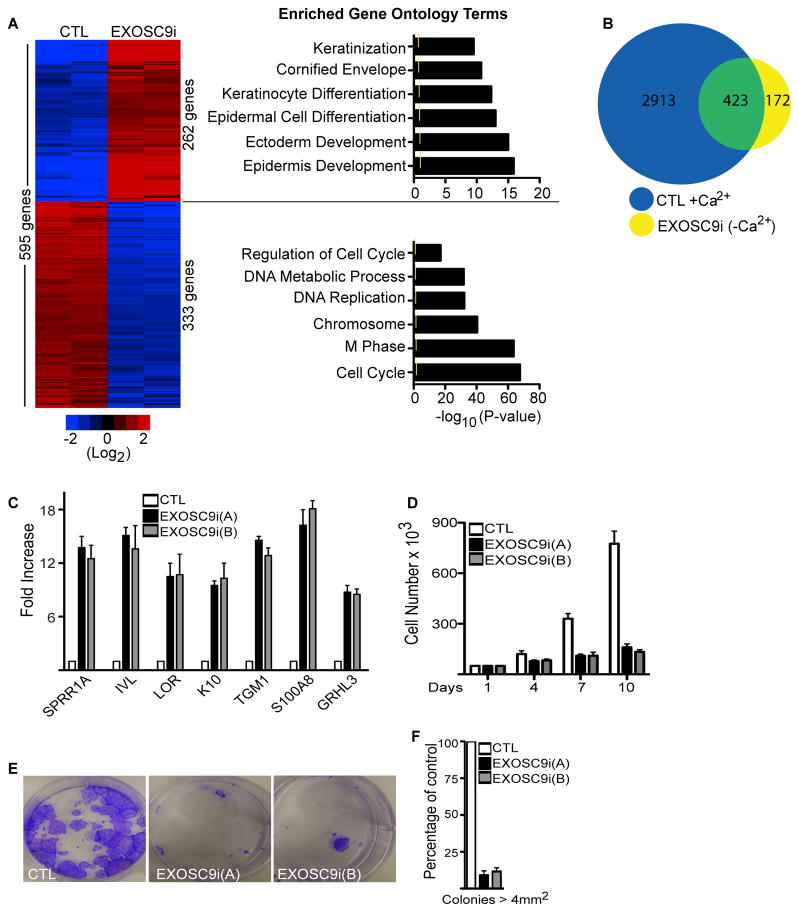
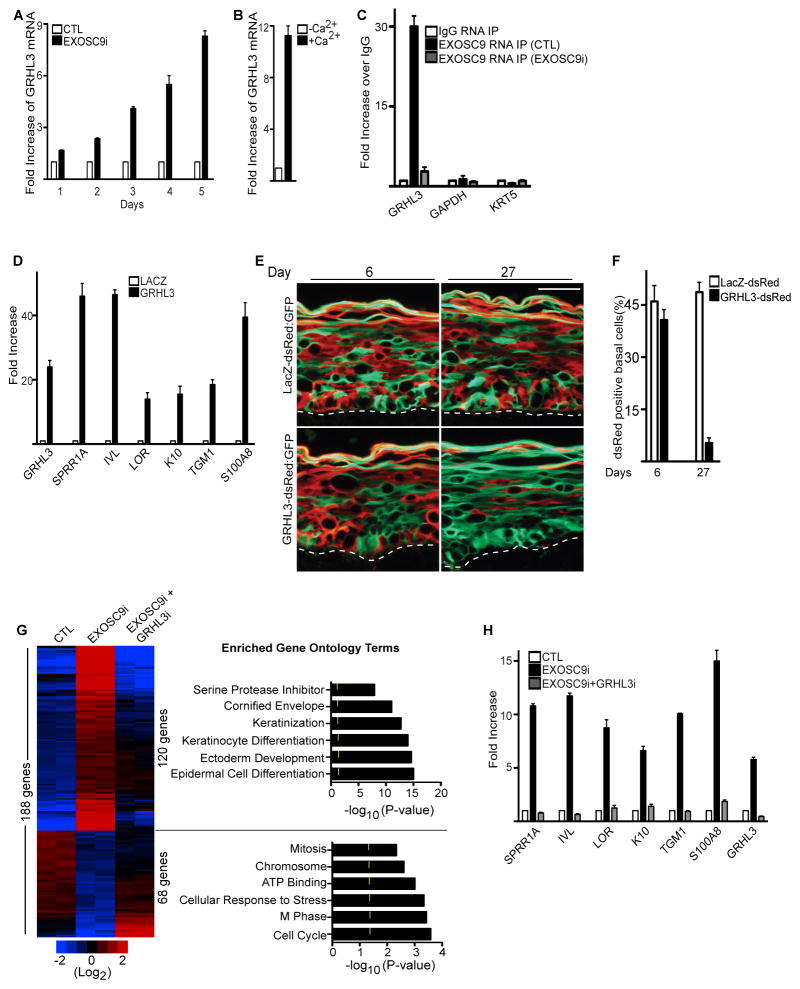
Stem and progenitor cells maintain the tissue they reside in for life by regulating the balance between proliferation and differentiation. How this is done is not well understood. Here, we report that the human exosome maintains progenitor cell function. The expression of several subunits of the exosome were found to be enriched in epidermal progenitor cells, which were required to retain proliferative capacity and to prevent premature differentiation. Loss of PM/Scl-75 also known as EXOSC9, a key subunit of the exosome complex, resulted in loss of cells from the progenitor cell compartment, premature differentiation, and loss of epidermal tissue. EXOSC9 promotes self-renewal and prevents premature differentiation by maintaining transcript levels of a transcription factor necessary for epidermal differentiation, GRHL3, at low levels through mRNA degradation. These data demonstrate that control of differentiation specific transcription factors through mRNA degradation is required for progenitor cell maintenance in mammalian tissue.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Andrulis ED, Werner J, Nazarian A, Erdjument-Bromage H, Tempst P, Lis JT. The RNA processing exosome is linked to elongating RNA polymerase II in Drosophila. Nature. 2002;420:837–841. - PubMed
-
- Belostotsky D. Exosome complex and pervasive transcription in eukaryotic genomes. Curr Opin Cell Biol. 2009;21:352–358. - PubMed
-
- Belostotsky DA, Sieburth LE. Kill the messenger: mRNA decay and plant development. Curr Opin Plant Biol. 2009;12:96–102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
