

Layer 6 cortical neurons require Reelin-Dab1 signaling for cellular orientation, Golgi deployment, and directed neurite growth into the marginal zone
- PMID: 22770513
- PMCID: PMC3466444
- DOI: 10.1186/1749-8104-7-25
Layer 6 cortical neurons require Reelin-Dab1 signaling for cellular orientation, Golgi deployment, and directed neurite growth into the marginal zone
Abstract
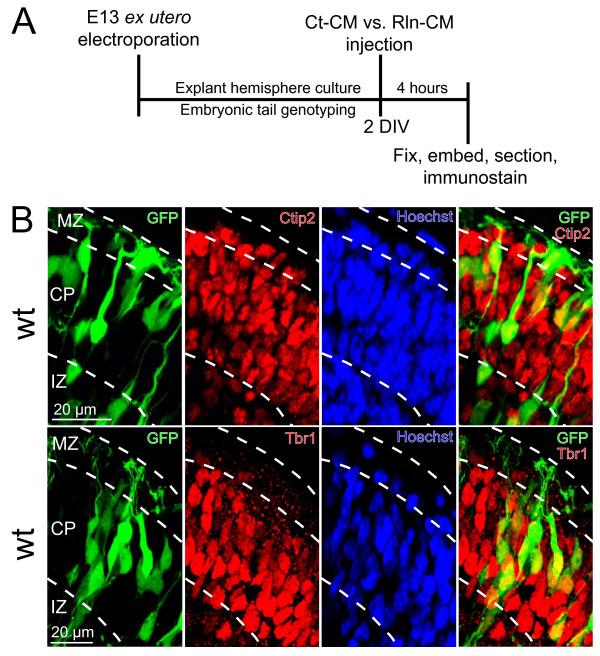
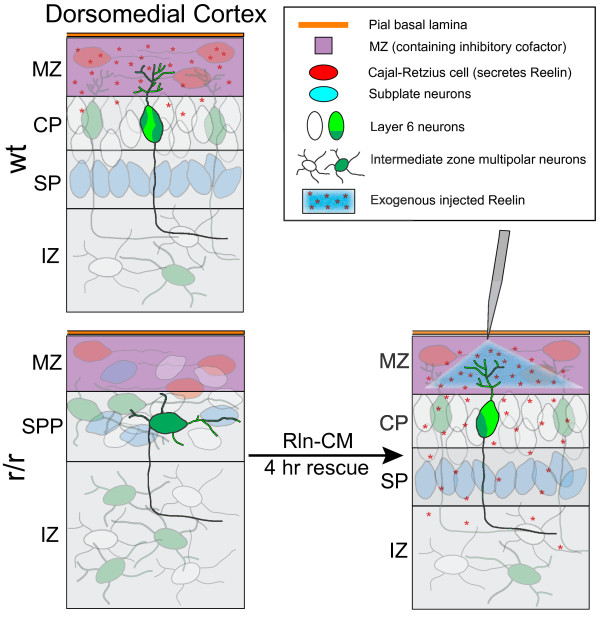
Background: The secreted ligand Reelin is believed to regulate the translocation of prospective layer 6 (L6) neocortical neurons into the preplate, a loose layer of pioneer neurons that overlies the ventricular zone. Recent studies have also suggested that Reelin controls neuronal orientation and polarized dendritic growth during this period of early cortical development. To explicitly characterize and quantify how Reelin controls this critical aspect of neurite initiation and growth we used a new ex utero explant model of early cortical development to selectively label a subset of L6 cortical neurons for complete 3-D reconstruction.
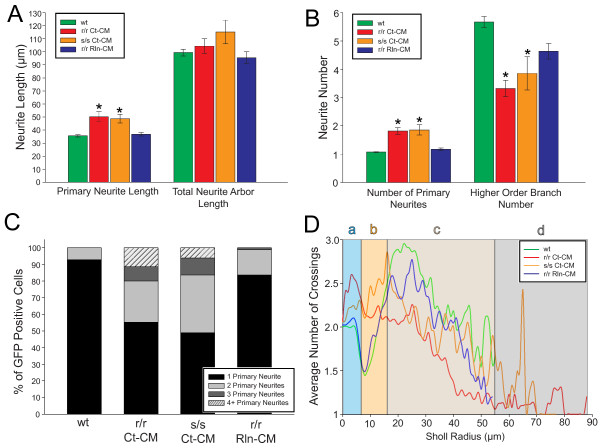
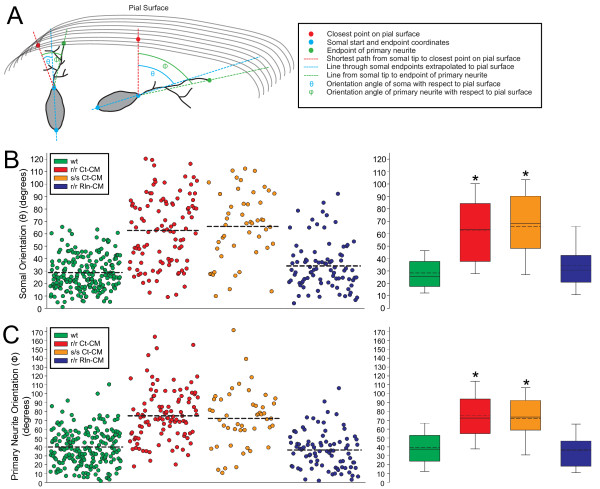
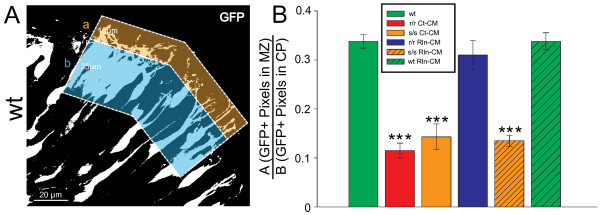
Results: The total neurite arbor sizes of neurons in Reelin-deficient (reeler mutant) and Dab1-deficient (Reelin-non-responsive scrambler mutant) cortices were quantified and unexpectedly were not different than control arbor lengths (p = 0.51). For each mutant, however, arbor organization was markedly different: mutant neurons manifested more primary processes (neurites emitted directly from the soma) than wild type, and these neurites were longer and displayed less branching. Reeler and scrambler mutant neurites extended tangentially rather than radially, and the Golgi apparatus that normally invests the apical neurite was compact in both reeler and scrambler mutants. Mutant cortices also exhibited a neurite "exclusion zone" which was relatively devoid of L6 neuron neurites and extended at least 15 μm beneath the pial surface, an area corresponding to the marginal zone (MZ) in the wild type explants. The presence of an exclusion zone was also indicated in the orientation of mutant primary neurite and neuronal somata, which failed to adopt angles within ~20˚ of the radial line to the pial surface. Injection of recombinant Reelin to reeler, but not scrambler, mutant cortices fully rescued soma orientation, Golgi organization, and dendritic projection defects within four hrs.
Conclusions: These findings indicate Reelin promotes directional dendritic growth into the MZ, an otherwise exclusionary zone for L6 neurites.
Figures


Similar articles
-
Reelin Prevents Apical Neurite Retraction during Terminal Translocation and Dendrite Initiation.J Neurosci. 2015 Jul 29;35(30):10659-74. doi: 10.1523/JNEUROSCI.1629-15.2015. J Neurosci. 2015. PMID: 26224852 Free PMC article.
-
Reelin promotes neuronal orientation and dendritogenesis during preplate splitting.Cereb Cortex. 2010 Sep;20(9):2213-23. doi: 10.1093/cercor/bhp303. Epub 2010 Jan 11. Cereb Cortex. 2010. PMID: 20064940 Free PMC article.
-
Altered speeds and trajectories of neurons migrating in the ventricular and subventricular zones of the reeler neocortex.Cereb Cortex. 2011 May;21(5):1018-27. doi: 10.1093/cercor/bhq168. Epub 2010 Sep 16. Cereb Cortex. 2011. PMID: 20847150
-
[Cytoarchitectonic abnormality in the facial nucleus of the reeler mouse].Kaibogaku Zasshi. 1999 Aug;74(4):411-20. Kaibogaku Zasshi. 1999. PMID: 10496086 Review. Japanese.
-
Reelin deficiency causes granule cell dispersion in epilepsy.Exp Brain Res. 2010 Jan;200(2):141-9. doi: 10.1007/s00221-009-1948-5. Epub 2009 Jul 26. Exp Brain Res. 2010. PMID: 19633980 Review.
Cited by
-
Reelin Prevents Apical Neurite Retraction during Terminal Translocation and Dendrite Initiation.J Neurosci. 2015 Jul 29;35(30):10659-74. doi: 10.1523/JNEUROSCI.1629-15.2015. J Neurosci. 2015. PMID: 26224852 Free PMC article.
-
Delayed cortical development in mice with a neural specific deletion of β1 integrin.Front Neurosci. 2023 May 12;17:1158419. doi: 10.3389/fnins.2023.1158419. eCollection 2023. Front Neurosci. 2023. PMID: 37250402 Free PMC article.
-
GRASPs link Reelin to the Golgi during neocortical development to control neuronal migration and dendritogenesis.Commun Biol. 2025 Apr 6;8(1):572. doi: 10.1038/s42003-025-08014-x. Commun Biol. 2025. PMID: 40188221 Free PMC article.
-
Ex utero electroporation and whole hemisphere explants: a simple experimental method for studies of early cortical development.J Vis Exp. 2013 Apr 3;(74):50271. doi: 10.3791/50271. J Vis Exp. 2013. PMID: 23609059 Free PMC article.
-
Reelin Counteracts Chondroitin Sulfate Proteoglycan-Mediated Cortical Dendrite Growth Inhibition.eNeuro. 2020 Jul 28;7(4):ENEURO.0168-20.2020. doi: 10.1523/ENEURO.0168-20.2020. Print 2020 Jul/Aug. eNeuro. 2020. PMID: 32641498 Free PMC article.
References
-
- Dotti CG, Banker GA. Experimentally induced alteration in the polarity of developing neurons. Nature. 1987;330:254–256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
