

Ultraviolet light provides a major input to non-image-forming light detection in mice
- PMID: 22771039
- PMCID: PMC3414846
- DOI: 10.1016/j.cub.2012.05.032
Ultraviolet light provides a major input to non-image-forming light detection in mice
Abstract
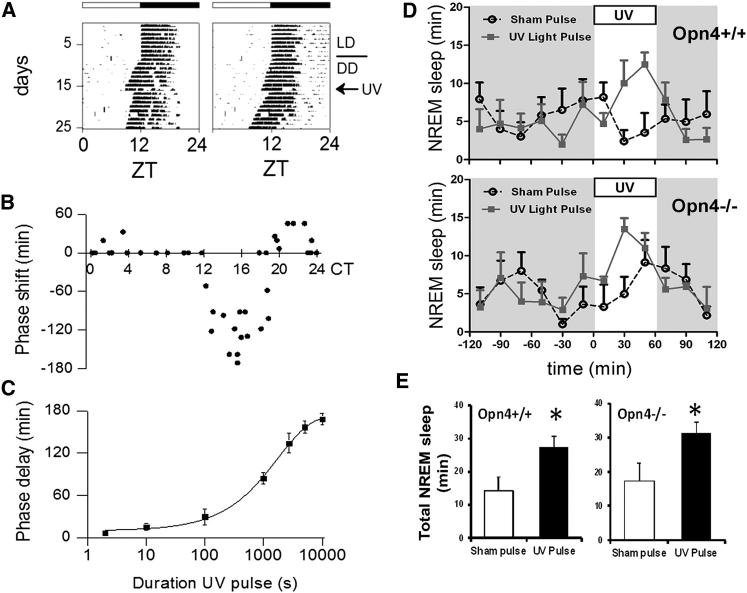
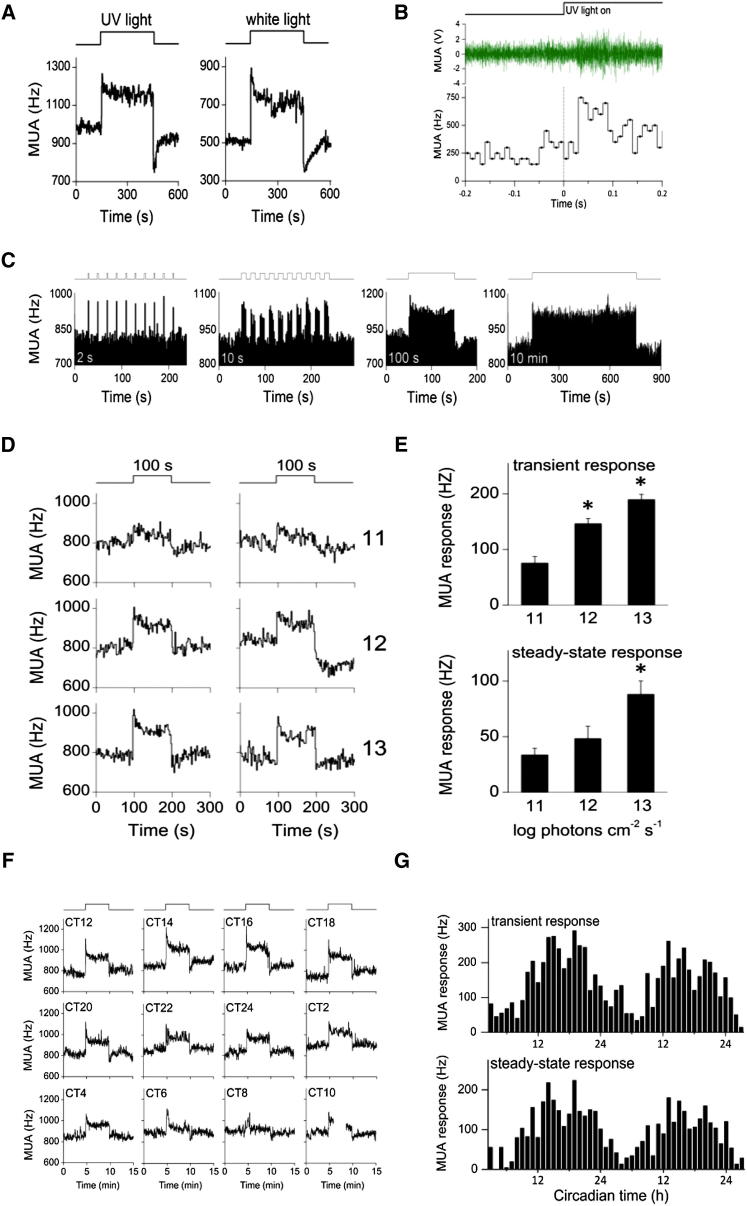
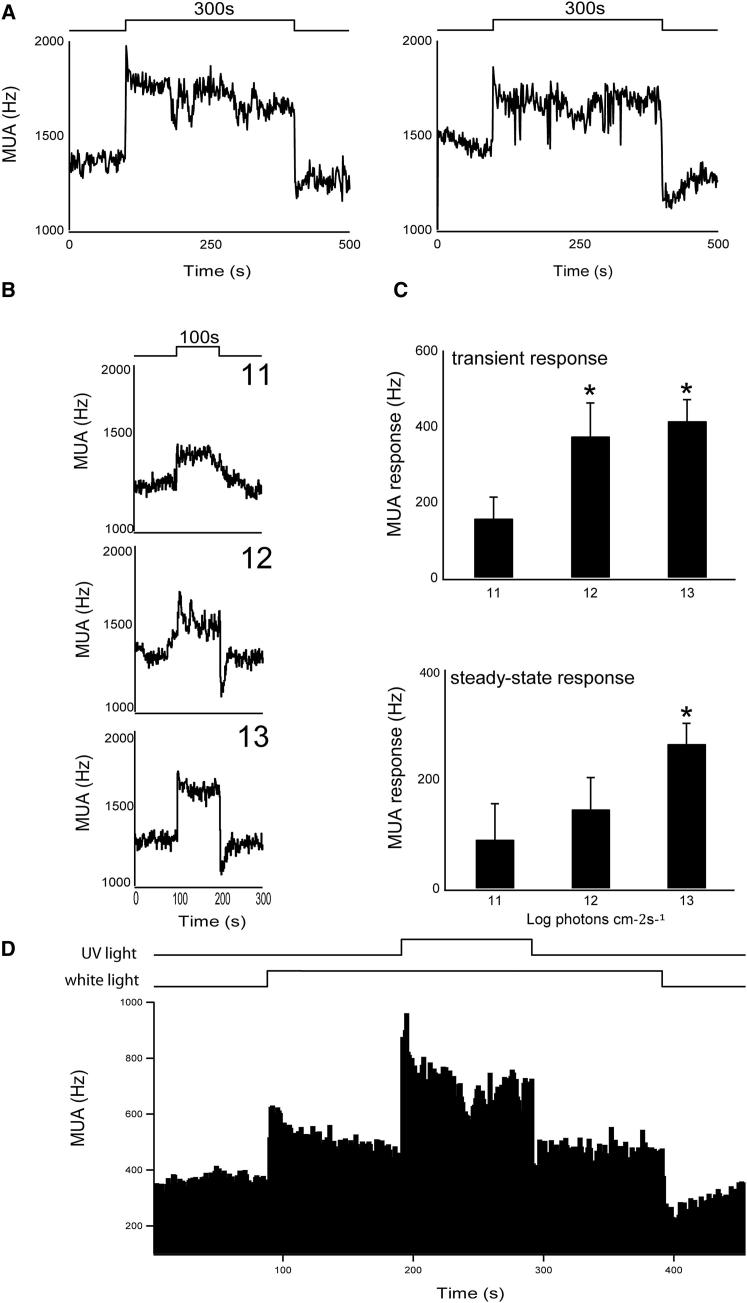
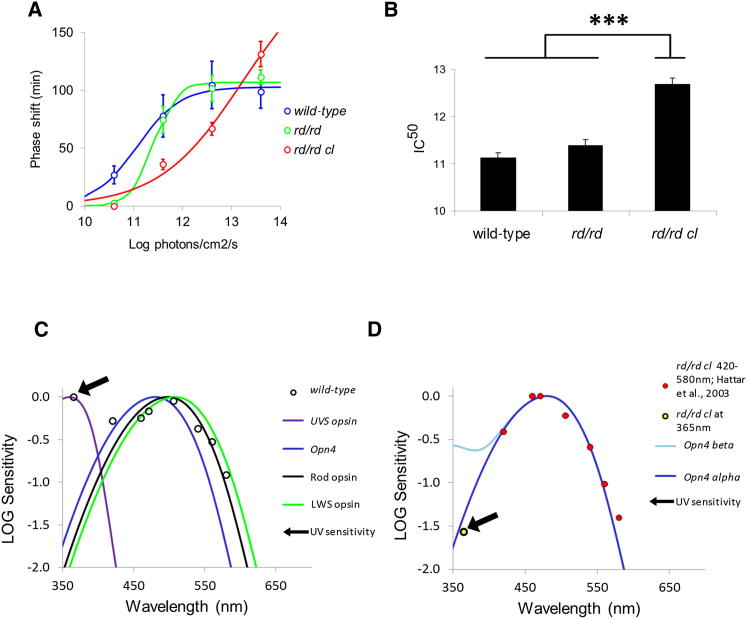
The change in irradiance at dawn and dusk provides the primary cue for the entrainment of the mammalian circadian pacemaker. Irradiance detection has been ascribed largely to melanopsin-based phototransduction [1-5]. Here we examine the role of ultraviolet-sensitive (UVS) cones in the modulation of circadian behavior, sleep, and suprachiasmatic nucleus (SCN) electrical activity. UV light exposure leads to phase-shifting responses comparable to those of white light. Moreover, UV light exposure induces sleep in wild-type and melanopsin-deficient (Opn4(-/-)) mice with equal efficacy. Electrical recordings from the SCN of wild-type mice show that UV light elicits irradiance-dependent sustained responses that are similar to those induced by white light, with characteristic fast transient components occurring at the light transitions. These responses are retained in Opn4(-/-) mice and preserved under saturating photopic conditions. The sensitivity of phase-shifting responses to UV light is unaffected by the loss of rods but is severely attenuated by the additional loss of cones. Our data show that UVS cones play an important role in circadian and sleep regulation in mice.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Berson D.M., Dunn F.A., Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295:1070–1073. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
