

Snail regulates MyoD binding-site occupancy to direct enhancer switching and differentiation-specific transcription in myogenesis
- PMID: 22771117
- PMCID: PMC4580277
- DOI: 10.1016/j.molcel.2012.05.046
Snail regulates MyoD binding-site occupancy to direct enhancer switching and differentiation-specific transcription in myogenesis
Abstract
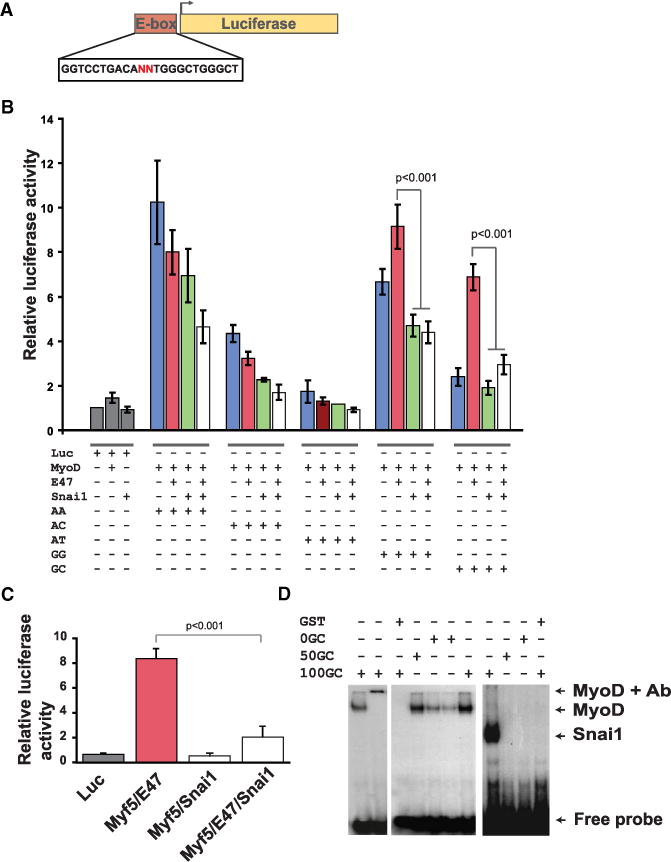
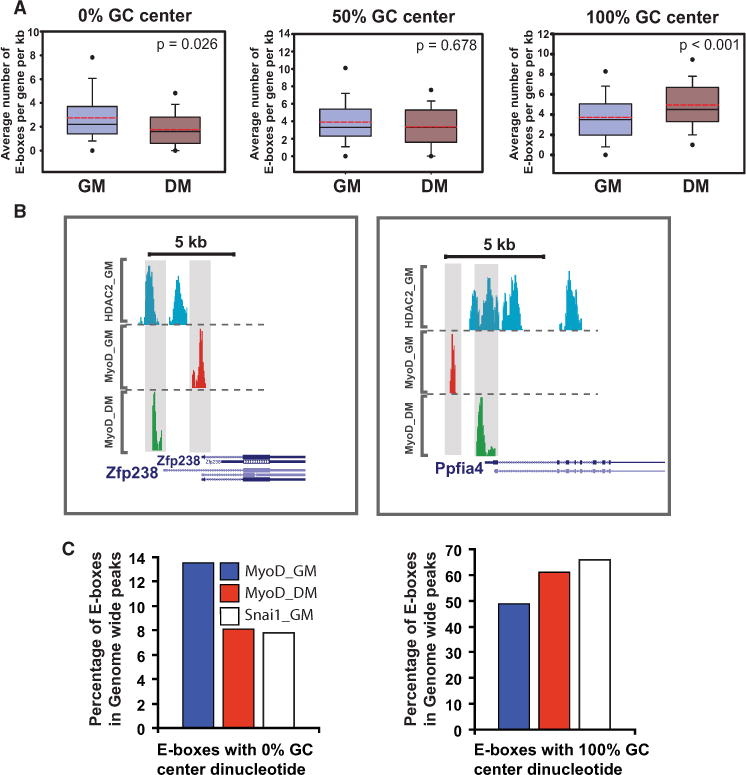
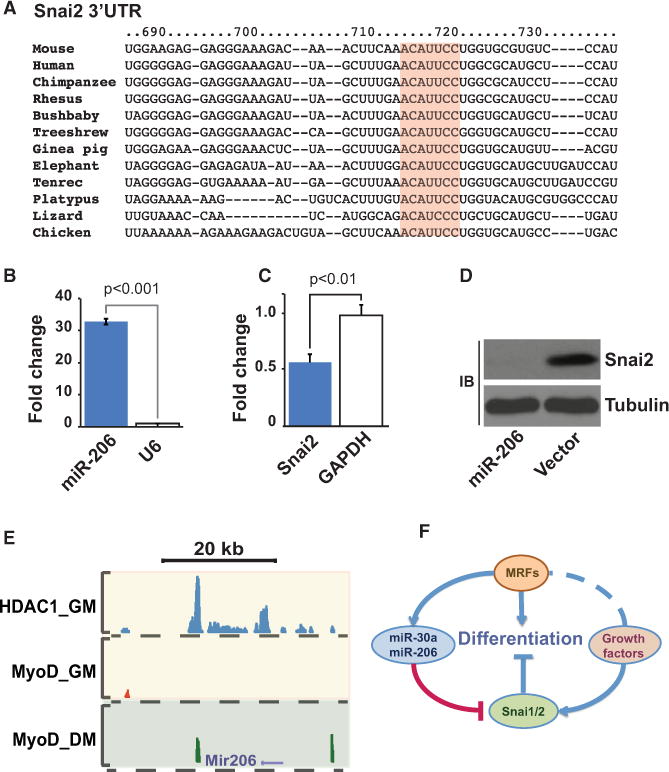
In skeletal myogenesis, the transcription factor MyoD activates distinct transcriptional programs in progenitors compared to terminally differentiated cells. Using ChIP-Seq and gene expression analyses, we show that in primary myoblasts, Snail-HDAC1/2 repressive complex binds and excludes MyoD from its targets. Notably, Snail binds E box motifs that are G/C rich in their central dinucleotides, and such sites are almost exclusively associated with genes expressed during differentiation. By contrast, Snail does not bind the A/T-rich E boxes associated with MyoD targets in myoblasts. Thus, Snai1-HDAC1/2 prevent MyoD occupancy on differentiation-specific regulatory elements, and the change from Snail to MyoD binding often results in enhancer switching during differentiation. Furthermore, we show that a regulatory network involving myogenic regulatory factors (MRFs), Snai1/2, miR-30a, and miR-206 acts as a molecular switch that controls entry into myogenic differentiation. Together, these results reveal a regulatory paradigm that directs distinct gene expression programs in progenitors versus terminally differentiated cells.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Barndt R, Dai MF, Zhuang Y. A novel role for HEB downstream or parallel to the pre-TCR signaling pathway during alpha beta thymopoiesis. J Immunol. 1999;163:3331–3343. - PubMed
-
- Bentzinger CF, Wang YX, Rudnicki MA. Building muscle: molecular regulation of myogenesis. Cold Spring Harb. Perspect Biol. 2012;4 http://dx.doi.org/10.1101/cshperspect.a008342. - DOI - PMC - PubMed
-
- Bergstrom DA, Penn BH, Strand A, Perry RL, Rudnicki MA, Tapscott SJ. Promoter-specific regulation of MyoD binding and signal transduction cooperate to pattern gene expression. Mol Cell. 2002;9:587–600. - PubMed
-
- Berkes CA, Tapscott SJ. MyoD and the transcriptional control of myogenesis. Semin Cell Dev Biol. 2005;16:585–595. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
