

Reconciling views on T cell receptor germline bias for MHC
- PMID: 22771140
- PMCID: PMC3983780
- DOI: 10.1016/j.it.2012.05.005
Reconciling views on T cell receptor germline bias for MHC
Abstract
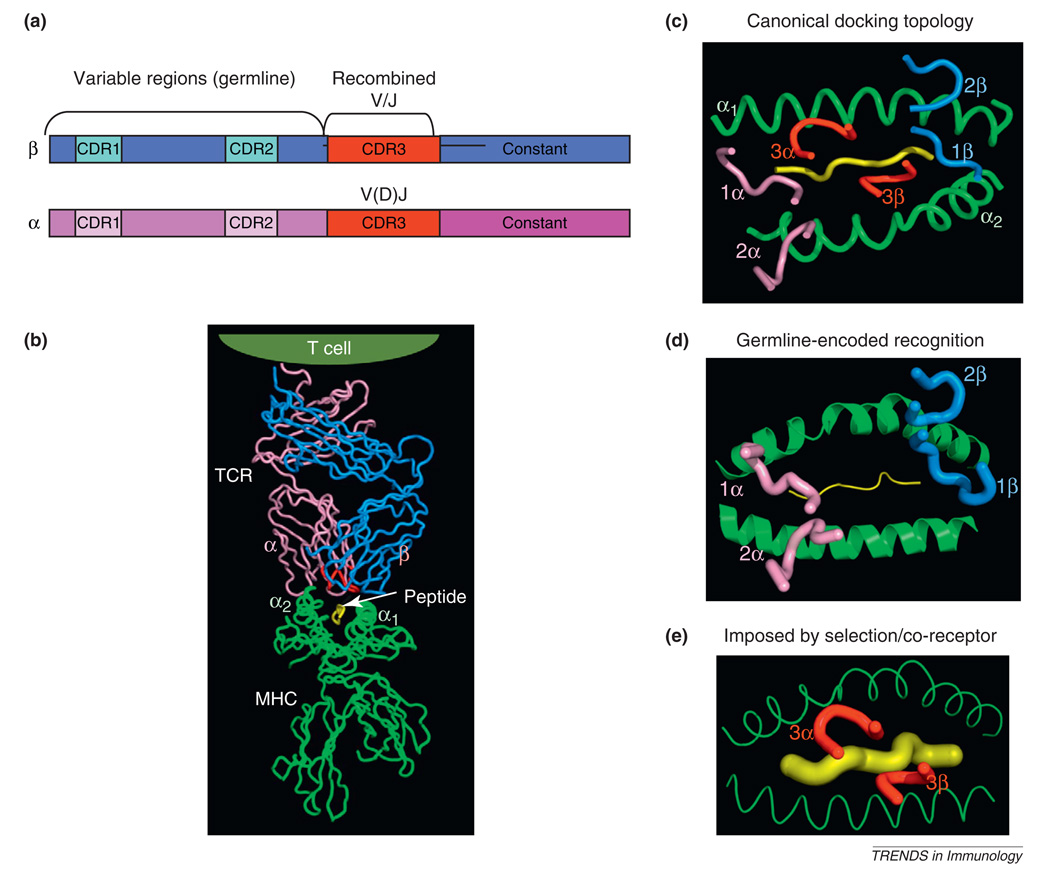
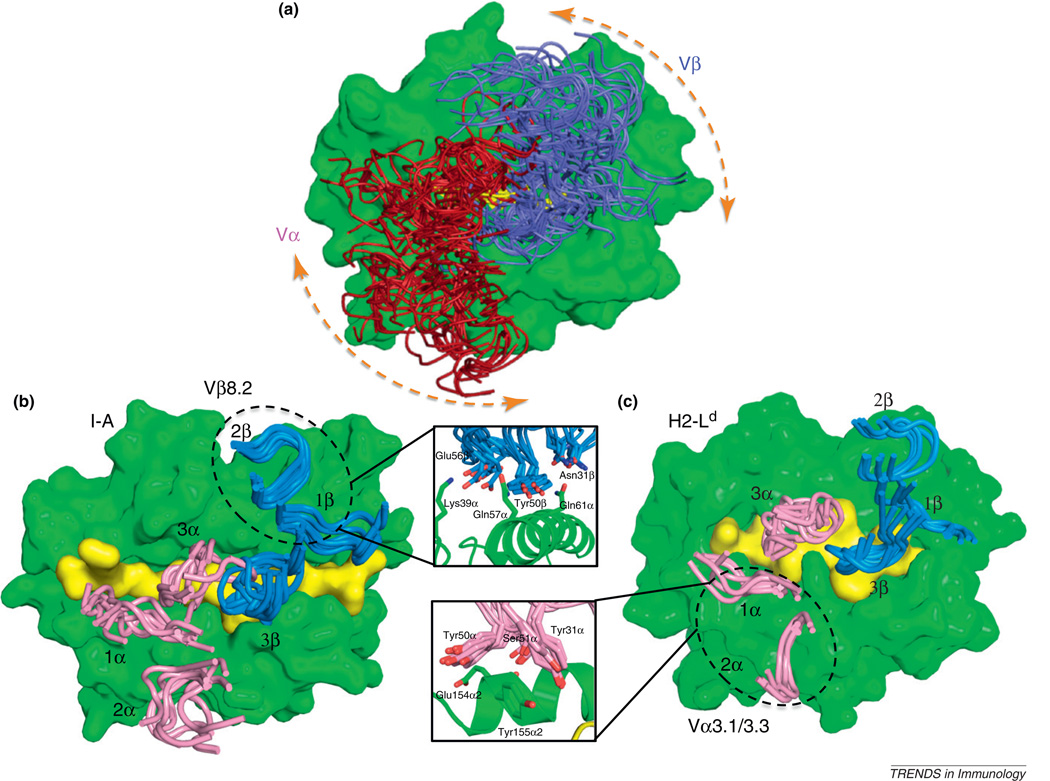
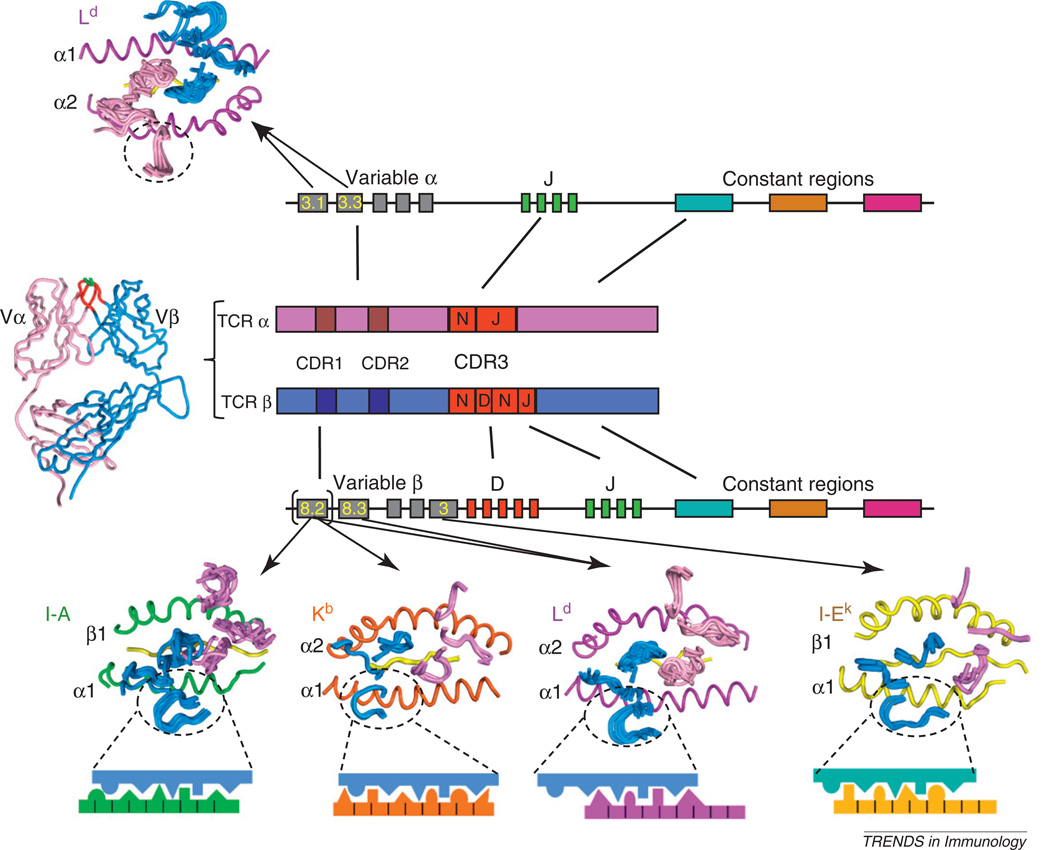
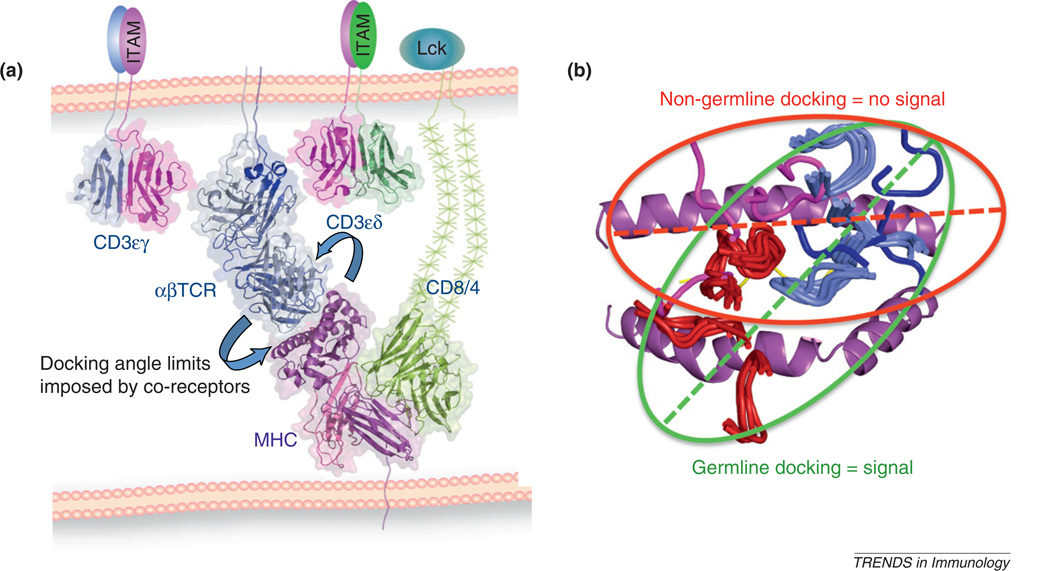
Whether MHC restriction by the T cell receptor (TCR) is a product of evolutionary pressures leading to germline-encoded 'rules of engagement' remains avidly debated. Structural results derived from analysis of TCR-peptide-MHC complexes appear to support a model of physical specificity between TCR germline V regions and MHC. Yet, some recent evidence suggests that thymic selection, and co-receptors may have misled us into thinking the TCR is exclusively MHC-specific, when in fact, TCRs can robustly engage non-MHC ligands when given the chance. Here, I propose that seemingly contradictory data and hypotheses for, and against, germline bias are, in fact, compatible and can be reconciled into a unifying model.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Huseby E, et al. TCR-MHC/peptide interactions: kissing-cousins or a shotgun wedding? Eur. J. Immunol. 2004;34:1243–1250. - PubMed
-
- Felix NJ, Allen PM. Specificity of T-cell alloreactivity. Nat. Rev. Immunol. 2007;7:942–953. - PubMed
-
- Germain RN. Immunology. Making a molecular match. Nature. 1990;344:19–22. - PubMed
-
- Collins EJ, Riddle DS. TCR-MHC docking orientation: natural selection, or thymic selection? Immunol. Res. 2008;41:267–294. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
