

Review
doi: 10.1007/s12576-012-0220-0.
Epub 2012 Jul 7.
Acidification of the synaptic cleft of cone photoreceptor terminal controls the amount of transmitter release, thereby forming the receptive field surround in the vertebrate retina
Affiliations
- PMID: 22773408
- PMCID: PMC10717482
- DOI: 10.1007/s12576-012-0220-0
Item in Clipboard
Review
Acidification of the synaptic cleft of cone photoreceptor terminal controls the amount of transmitter release, thereby forming the receptive field surround in the vertebrate retina
J Physiol Sci.
2012 Sep.
Abstract
In the vertebrate retina, feedback from horizontal cells (HCs) to cone photoreceptors plays a key role in the formation of the center-surround receptive field of retinal cells, which induces contrast enhancement of visual images. The mechanism underlying surround inhibition is not fully understood. In this review, we discuss this issue, focusing on our recent hypothesis that acidification of the synaptic cleft of the cone photoreceptor terminal causes this inhibition by modulating the Ca channel of the terminals. We present evidence that the acidification is caused by proton excretion from HCs by a vacuolar type H(+) pump. Recent publications supporting or opposing our hypothesis are discussed.
Figures

Surround illumination augments cone I
Ca. A
I
Ca of a cone in newt retinal slice was recorded under the whole-cell voltage clamp condition at a holding potential of −40 mV. The Cs+-based pipette solution contained 20 mM BAPTA to minimize any Ca2+-activated currents. Depolarizing voltage steps from −50 mV to +8 mV (2 mV step) induced inward currents. I
Ca of a cone in newt retinal slice was recorded under the whole-cell voltage clamp condition at a holding potential of −40 mV. The I
Ca was abolished by extracellular cadmium, a blocker of Ca channels (plots with open squares shown in the right panel). Five representative traces, voltage-clamped at −40, −26, −24, −16 and −4 mV, are shown. Diffuse illumination (4,000 μm in diameter used for ‘surround illumination’: shorter bar) was applied every 4 s, while the spot illumination (30 μm in diameter: top bar) was maintained. An additional 2-mV depolarization was applied to mimic an ephaptic (field) effect (external voltage drop) after withdrawing the diffuse illumination. Note that at −4 mV (pink trace), diffuse illumination evoked an inward current, while a +2-mV pulse evoked an outward current. The current amplitude was sampled at the time indicated by the symbols to construct the I–V curves shown in B
a and B
b. B
a Leak-subtracted I–V curve of cone I
Ca in the presence of the spot (filled squares) and during diffuse illumination (open squares). Inset shows activation curves fitted to the Boltzmann function derived from the I–V curves. B
b Leak-subtracted I–V curve of cone I
Ca in the presence of the spot light (filled circles) and during a +2-mV depolarizing pulse (open circles). Inset shows activation curves fitted to the Boltzmann function derived from the I–V curves. Boxed inset isolation method of I–V curves of cone I
Ca obtained from a different cone in A. Top The I–V curvers were obtained in the control solution [filled squares (1)] and in a 3 mM Cd containing solution [open squares (2)]. Bottom (open circles) I–V curve of the cone I
Ca obtained by subtracting the I–V curve in a 3 mM Cd containing solution from that in the control solution [(1)–(2)]. Filled circles
I–V curve obtained by subtracting the I–V curve from the extrapolated leakage current from that in the control solution [(1)–(3)] (from Hirasawa and Kaneko [28])

Modulation of cone I
Ca by a high-pH solution applied to the cone terminal layer. A Alkalinized Ringer’s solution (pH 9.0) was focally ejected to cone synaptic terminal layer in newt retinal slice. The cone was voltage-clamped at various voltages in the range of −50 to +6 mV in 8-mV steps. The representative four traces, voltage clamped at −42, −26, −18 and +6 mV, are shown. The current was sampled at the points marked by a symbol to construct the I–V curves shown in B. Small spot illumination (30 μm in diameter) was maintained throughout. B
Top leak-subtracted I–V curve of cone I
Ca in normal Ringer’s solution (pH 7.4, filled squares) and in response to a high-pH solution (pH 9.0, open circles). Bottom activation curves derived from the I–V curves fitted to the Boltzmann function (from Hirasawa and Kaneko [28])
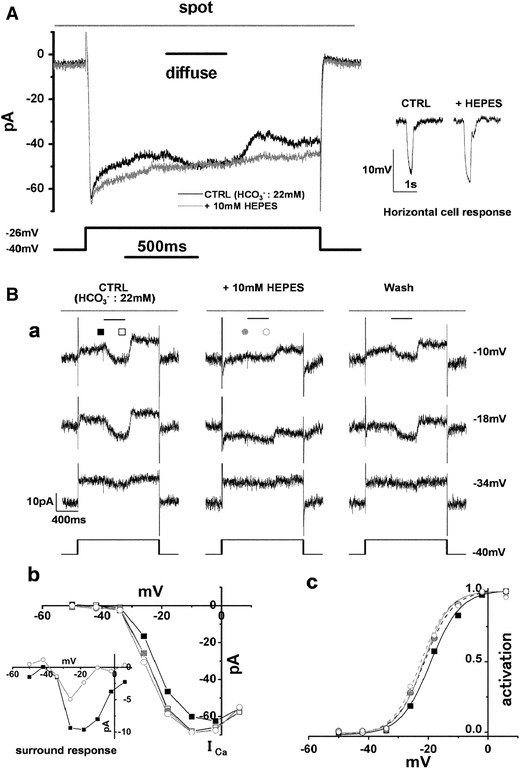
Cone I
Ca and its surround response recorded in a superfusate enriched with HEPES. A Effects of 10 mM HEPES-enriched buffer on cone I
Ca and surround response. The cone in newt retinal slice was depolarized from the holding voltage of −40 to −26 mV. Diffuse light illumination (4,000 μm in diameter used for surround illumination: shorter bar) was given during the step depolarization in the presence of a small spot light (30 μm in diameter: top bar). Changing the superfusate from control solution (black trace) to one supplemented with 10 mM HEPES increased cone I
Ca and prevented any additional inward current upon surround illumination (gray trace). Inset shows horizontal cell (HC) responses to a large light spot (4,000 μm in diameter) in the control solution and in the solution enriched with HEPES. B
a Reversible effects of 10 mM HEPES-enriched buffer on cone I
Ca and surround response. The small spot light (30 μm in diameter: top bars) was kept on throughout. Diffuse light illumination (4,000 μm in diameter: shorter bar) was given during the step depolarization in the presence of the small spot. The cone was held at −40 mV and polarized to voltages ranging from −50 to +6 mV in 8 mV steps. The current traces before, during and after application of 10 mM HEPES-enriched buffer. The recording sequence was left column (−34, −18 and −10 mV) followed by the middle column and finally the right column. The leak conductance of 2.26 nS did not change either in the HEPES-containing solution or during the washout (current traces at −34 mV). In the HEPES-containing solution, inward I
Ca in darkness was reversibly increased, and the surround response was reversibly suppressed (current traces at −18 and −10 mV). Symbols denote the sampling points for calculation of the I–V curves of I
Ca in B
b. In the control solution, the inward current produced by I
Ca was counterbalanced by the outward leak current (at −18 and −10 mV in the control and washout solutions). B
b
I–V curves of cone I
Ca recorded in B
a. The leak conductance was subtracted. Filled and open squares in the control solution without and with surround illumination, respectively; filled and open gray circles in the HEPES-containing solution without and with surround illumination, respectively. Inset shows the voltage dependence of the surround response in the control solution (filled squares) and in the HEPES-containing solution (open circles). B
c Activation curves fitted to the Boltzmann function derived from the data in B
b (from Hirasawa and Kaneko [28])
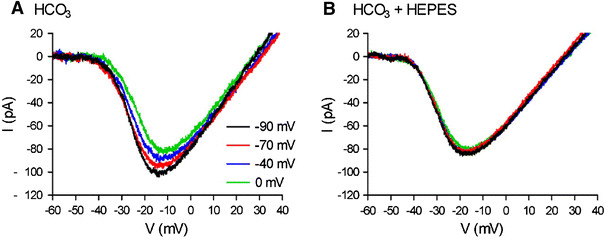
Changes in horizontal cell (HC) membrane potential alter the amplitude and voltage dependence of I
Ca in an adjacent cone simultaneously recorded: these effects are blocked by application of HEPES. A
I
Ca of cone in tiger salamander retinal slice was recorded under the whole-cell voltage clamp condition with a ramp-function from −90 to +60 mV (0.5 mV/ms) applied from a steady holding potential of −70 mV. By step-wise depolarization of HC simultaneously voltage-clamping at −90, −70, −40 and 0 mV, the cone I
Ca was progressively decreased in amplitude and positively shifted in the activation voltage to more positive potentials. B Addition of HEPES (10 mM) abolished the voltage shift and amplitude changes of cone I
Ca curves produced by the HC polarization (from Cadetti and Thoreson [55])
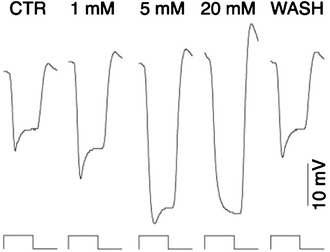
Concentration-dependent effects of HEPES on monkey HC light response. Light responses of an HC in macaque monkey retina illuminated to a 2,000-μm diameter spot light were obtained by intracellular recording. HEPES at 1, 5 and 20 mM increased the hyperpolarization amplitude and decreased the slow depolarization in a dose-dependent manner (from Davenport et al. [64])
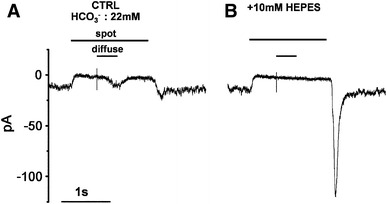
Center and surround responses of an OFF-type bipolar cell (BC) recorded in a solution enriched with HEPES. Light responses of an OFF-type BC in newt retinal slice were recorded under the voltage-clamp conditions (holding voltage, −40 mV). A In control Ringer’s solution, spot illumination (50 μm in diameter) evoked an outward current as a center response and surround illumination (diffuse light superimposed on the spot light: 4,000 μm in diameter) evoked an inward current as a surround response. B Changing the superfusate to the HEPES-enriched solution (10 mM) abolished the surround response. The transient inward current at the time of switch-off of spot illumination was prominently increased in HEPES buffer (from Hirasawa and Kaneko [28])

HEPES attenuates parasol ganglion cell (GC) surround. A An ON parasol GC in macaque monkey retina was illuminated to a 300-μm-diameter spot light (stimulus trace below) and its ON responses in the absence (left) and presence (right) of 20 mM HEPES were obtained by intracellular recording. The responses were similar in both conditions. B Responses of a parasol GC to a larger 2,000-μm-diameter spot (stimulus trace below) were obtained in the absence (left) and presence (right) of HEPES. The control response was small and transient because of surround antagonism. The response in HEPES was larger and more sustained, indicating diminished surround antagonism (from Davenport et al. [64])
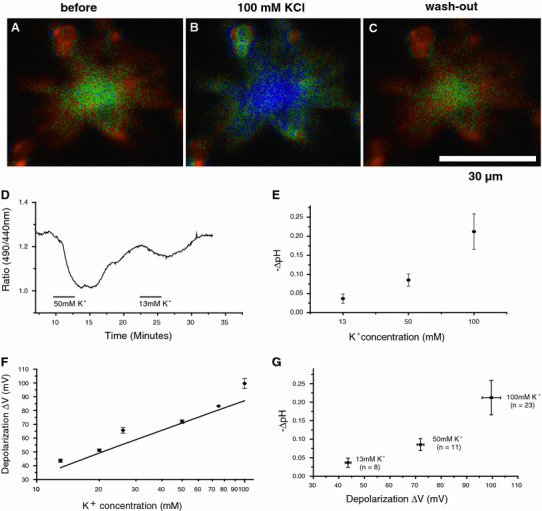
Acidification of extracellular surface of cone-driven HCs dissociated from goldfish retinas by high-K+-induced membrane depolarization. A–C A series of time-dependent surface pH (pHs) changes of an HC (pHs reduction: 0.10 pH units from 7.60 during 100 mM K+ application for 3 min). The pHs change is measured by the ratio imaging method (bluish and yellowish colour corresponds to the lower and higher pHs, respectively). D Acidification responses of the HC extracellular surface serially induced by 2 min bath applications of 50 and 13 mM KCl containing Ringers to depolarize the membrane potential. E Acidification of the measured HC extracellular surface is dependent on the KCl concentration. F HC membrane depolarization is dependent on the KCl concentration measured by patch-clamp study in current-clamp mode in comparison with the control Ringer having 2.6 mM KCl. Straight line Nernst equation for K+ shown by  . G HC acidification induced by high-K+ is dependent on the membrane depolarization. This graph was composed by combining E and F at 13, 50 and 100 mM KCl (from Jouhou et al. [29])
. G HC acidification induced by high-K+ is dependent on the membrane depolarization. This graph was composed by combining E and F at 13, 50 and 100 mM KCl (from Jouhou et al. [29])
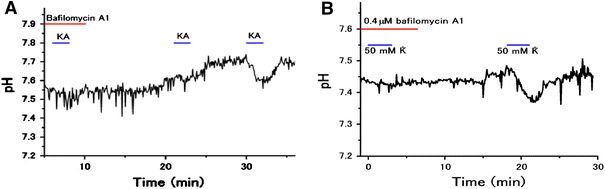
Bafilomycin A1, a specific inhibitor of V-ATPase inhibitor, blocks extracellular surface acidification of goldfish HCs induced by 20 μM kainate or 50 mM K+. A Kainate-induced acidification is blocked by 0.4 μM bafilomycin A1, and partially recovered after 10 min wash-out and then fully recovered after 20 min wash-out; B 0.4 μM bafilomycin A1 suppressed 50 mM K+-induced HC acidification, which was recovered after 13 min wash-out (from Jouhou et al. [29])
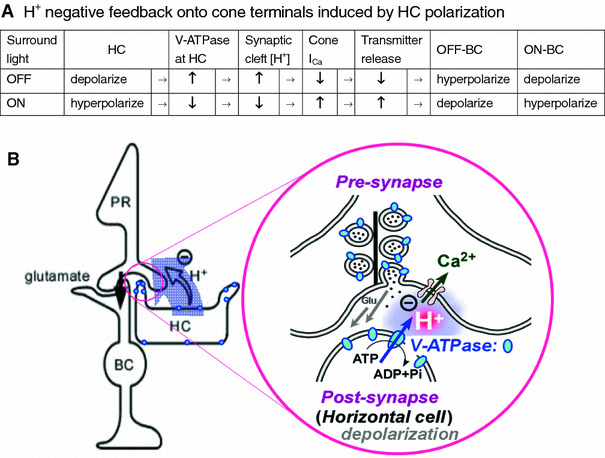
Schematics of H+ negative feedback onto BCs via H+ concentration change at synaptic clefts of cone terminals induced by HC polarization. A H+ negative feedback onto OFF- and ON-type BCs via the series of effects of depolarization or hyperpolarization of HCs. The depolarization or hyperpolarization of HCs is induced either by OFF or ON of light illumination, respectively. ↑ denotes an activation or increase, whereas ↓ denotes an inactivation or decrease. In the presence of HEPES, synaptic cleft pH changes will be suppressed and bring about a change to the opposite direction to the control effect of HC polarization. These effects of HEPES imply the suppression of negative feedback ordinarily present in the control condition. B A schematic drawing of H+ negative feedback onto BCs via the concentration increase of H+ released from depolarized HCs into the synaptic cleft of cone terminals, illustrating OFF case in A. H+ release is due to V-ATPase (a proton pump located on plasma membrane shown by blue circles in an expanded figure) on depolarized HCs, resulting in suppression of glutamate release from cones. V-ATPase is also localized at the membrane of synaptic vesicles at the cone terminals
Similar articles
-
pH changes in the invaginating synaptic cleft mediate feedback from horizontal cells to cone photoreceptors by modulating Ca2+ channels.J Gen Physiol. 2003 Dec;122(6):657-71. doi: 10.1085/jgp.200308863. Epub 2003 Nov 10. J Gen Physiol. 2003. PMID: 14610018 Free PMC article.
-
Proton-mediated feedback inhibition of presynaptic calcium channels at the cone photoreceptor synapse.J Neurosci. 2005 Apr 20;25(16):4108-17. doi: 10.1523/JNEUROSCI.5253-04.2005. J Neurosci. 2005. PMID: 15843613 Free PMC article.
-
Sources of protons and a role for bicarbonate in inhibitory feedback from horizontal cells to cones in Ambystoma tigrinum retina.J Physiol. 2016 Nov 15;594(22):6661-6677. doi: 10.1113/JP272533. Epub 2016 Jul 21. J Physiol. 2016. PMID: 27345444 Free PMC article.
-
The feedback pathway from horizontal cells to cones. A mini review with a look ahead.Vision Res. 1999 Jul;39(15):2449-68. doi: 10.1016/s0042-6989(99)00043-7. Vision Res. 1999. PMID: 10396615 Review.
-
Horizontal Cell Feedback to Cone Photoreceptors in Mammalian Retina: Novel Insights From the GABA-pH Hybrid Model.Front Cell Neurosci. 2020 Nov 4;14:595064. doi: 10.3389/fncel.2020.595064. eCollection 2020. Front Cell Neurosci. 2020. PMID: 33328894 Free PMC article. Review.
Cited by
-
Versatile functional roles of horizontal cells in the retinal circuit.Sci Rep. 2017 Jul 17;7(1):5540. doi: 10.1038/s41598-017-05543-2. Sci Rep. 2017. PMID: 28717219 Free PMC article.
-
Voltage-Gated Calcium Channels: Key Players in Sensory Coding in the Retina and the Inner Ear.Physiol Rev. 2018 Oct 1;98(4):2063-2096. doi: 10.1152/physrev.00030.2017. Physiol Rev. 2018. PMID: 30067155 Free PMC article. Review.
-
Specialized synaptic pathway for chromatic signals beneath S-cone photoreceptors is common to human, Old and New World primates.J Opt Soc Am A Opt Image Sci Vis. 2014 Apr 1;31(4):A189-94. doi: 10.1364/JOSAA.31.00A189. J Opt Soc Am A Opt Image Sci Vis. 2014. PMID: 24695169 Free PMC article.
-
Imaging an optogenetic pH sensor reveals that protons mediate lateral inhibition in the retina.Nat Neurosci. 2014 Feb;17(2):262-8. doi: 10.1038/nn.3627. Epub 2014 Jan 19. Nat Neurosci. 2014. PMID: 24441679 Free PMC article.
-
Activation of retinal glial (Müller) cells by extracellular ATP induces pronounced increases in extracellular H+ flux.PLoS One. 2018 Feb 21;13(2):e0190893. doi: 10.1371/journal.pone.0190893. eCollection 2018. PLoS One. 2018. PMID: 29466379 Free PMC article.
References
-
- Kuffler SW. Discharge patterns and functional organization of mammalian retina. J Neurophysiol. 1953;16:37–68. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
