

A flavin binding cryptochrome photoreceptor responds to both blue and red light in Chlamydomonas reinhardtii
- PMID: 22773746
- PMCID: PMC3426128
- DOI: 10.1105/tpc.112.098947
A flavin binding cryptochrome photoreceptor responds to both blue and red light in Chlamydomonas reinhardtii
Abstract
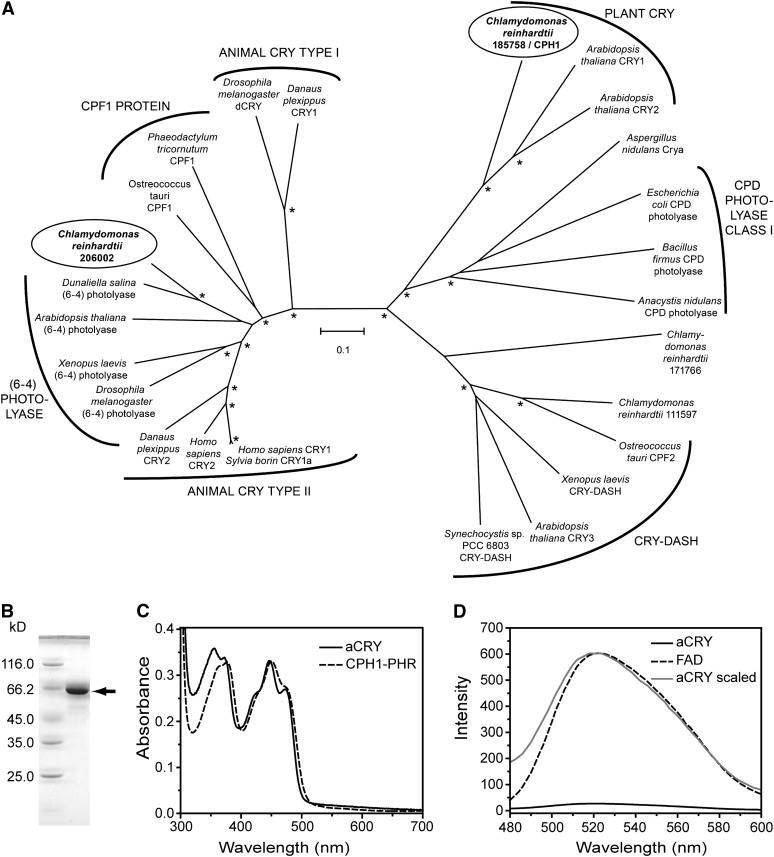
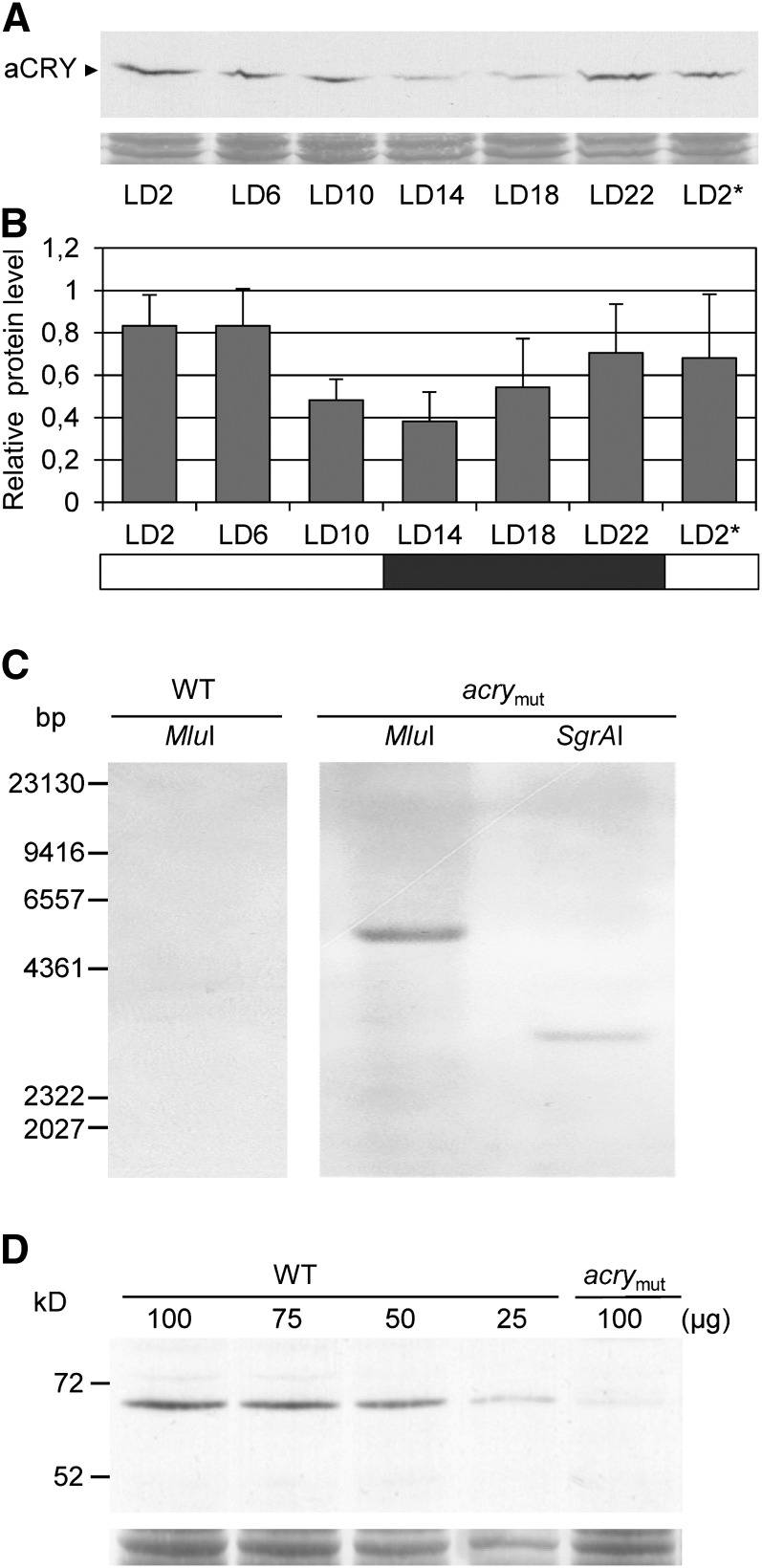
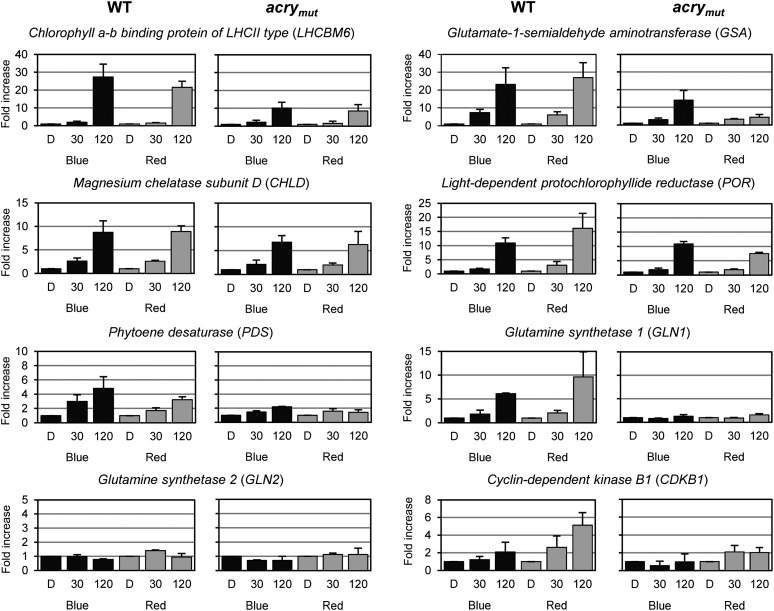
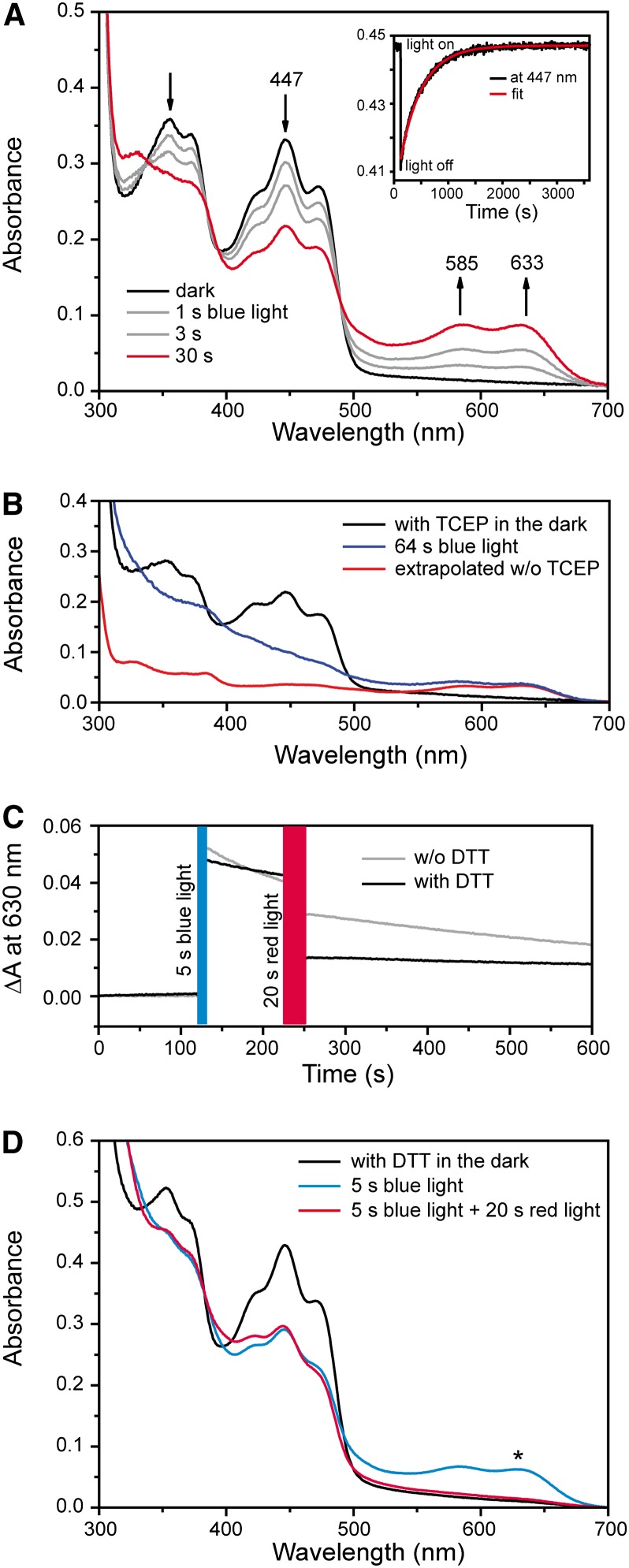
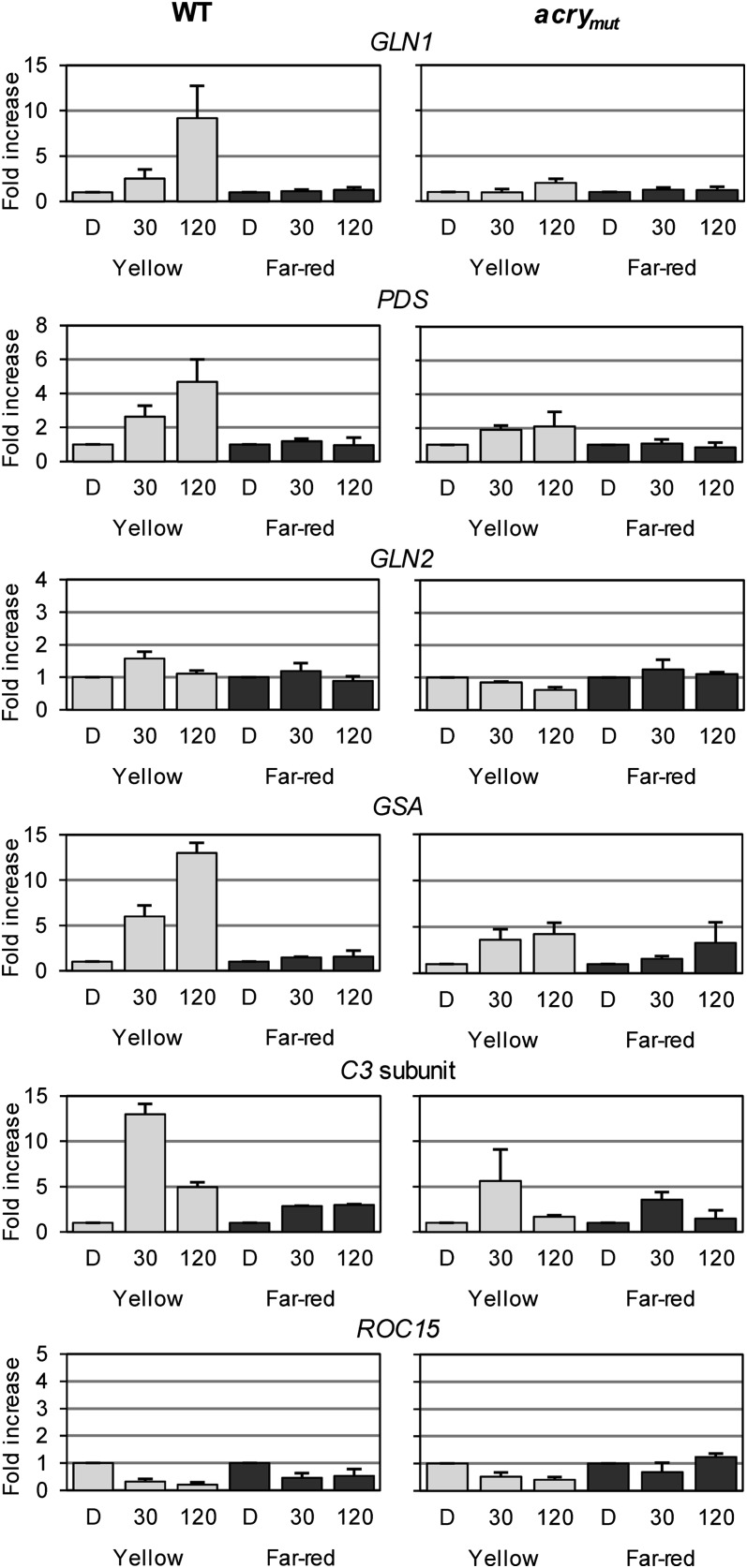
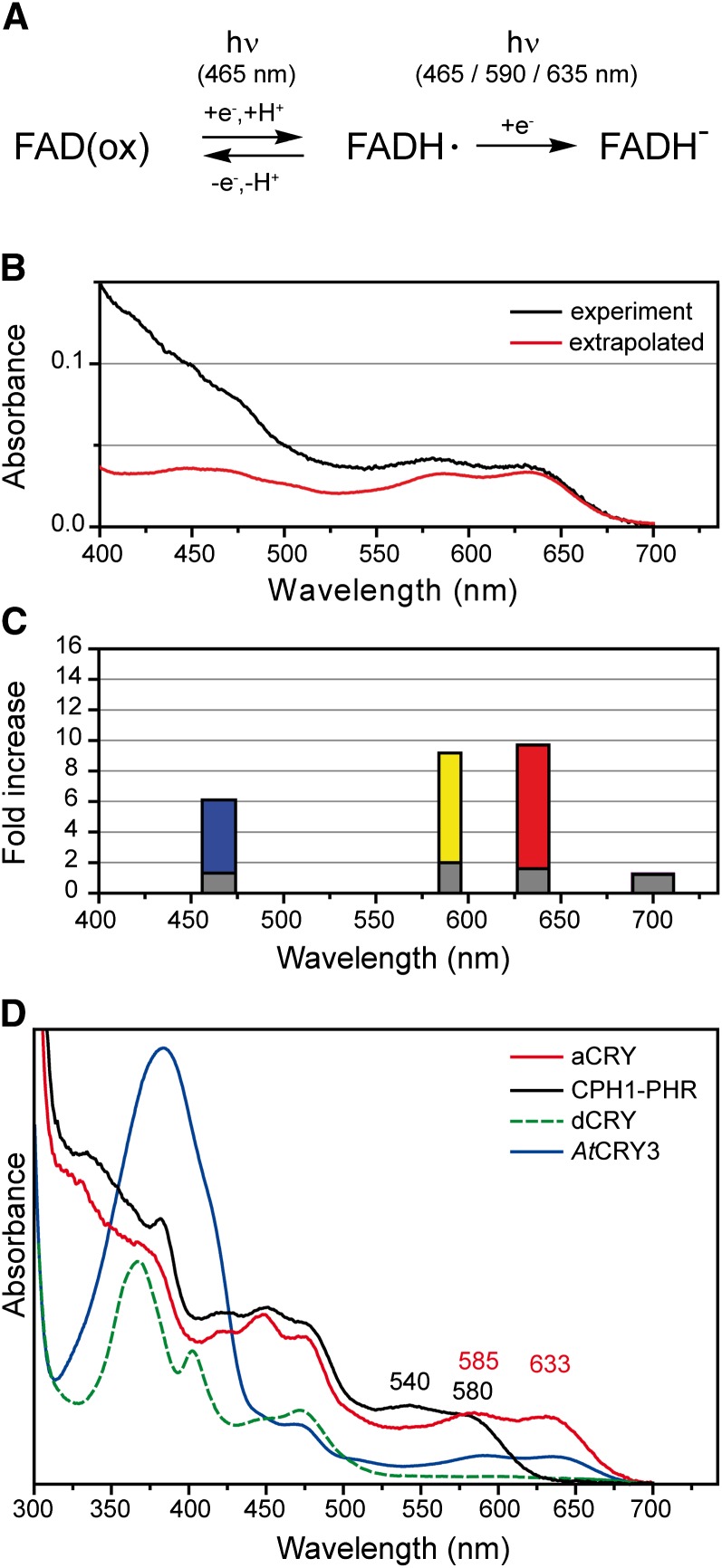
Cryptochromes are flavoproteins that act as sensory blue light receptors in insects, plants, fungi, and bacteria. We have investigated a cryptochrome from the green alga Chlamydomonas reinhardtii with sequence homology to animal cryptochromes and (6-4) photolyases. In response to blue and red light exposure, this animal-like cryptochrome (aCRY) alters the light-dependent expression of various genes encoding proteins involved in chlorophyll and carotenoid biosynthesis, light-harvesting complexes, nitrogen metabolism, cell cycle control, and the circadian clock. Additionally, exposure to yellow but not far-red light leads to comparable increases in the expression of specific genes; this expression is significantly reduced in an acry insertional mutant. These in vivo effects are congruent with in vitro data showing that blue, yellow, and red light, but not far-red light, are absorbed by the neutral radical state of flavin in aCRY. The aCRY neutral radical is formed following blue light absorption of the oxidized flavin. Red illumination leads to conversion to the fully reduced state. Our data suggest that aCRY is a functionally important blue and red light-activated flavoprotein. The broad spectral response implies that the neutral radical state functions as a dark form in aCRY and expands the paradigm of flavoproteins and cryptochromes as blue light sensors to include other light qualities.
Figures

Similar articles
-
News about cryptochrome photoreceptors in algae.Plant Signal Behav. 2013 Feb;8(2):e22870. doi: 10.4161/psb.22870. Epub 2012 Nov 15. Plant Signal Behav. 2013. PMID: 23154511 Free PMC article.
-
Essential Role of an Unusually Long-lived Tyrosyl Radical in the Response to Red Light of the Animal-like Cryptochrome aCRY.J Biol Chem. 2016 Jul 1;291(27):14062-14071. doi: 10.1074/jbc.M116.726976. Epub 2016 May 9. J Biol Chem. 2016. PMID: 27189948 Free PMC article.
-
Response of the Sensory animal-like cryptochrome aCRY to blue and red light as revealed by infrared difference spectroscopy.Biochemistry. 2014 Feb 18;53(6):1041-50. doi: 10.1021/bi401599z. Epub 2014 Feb 5. Biochemistry. 2014. PMID: 24467183
-
Cryptochrome photoreceptors in green algae: Unexpected versatility of mechanisms and functions.J Plant Physiol. 2017 Oct;217:4-14. doi: 10.1016/j.jplph.2017.05.021. Epub 2017 May 31. J Plant Physiol. 2017. PMID: 28619534 Review.
-
Cryptochromes and the Circadian Clock: The Story of a Very Complex Relationship in a Spinning World.Genes (Basel). 2021 Apr 29;12(5):672. doi: 10.3390/genes12050672. Genes (Basel). 2021. PMID: 33946956 Free PMC article. Review.
Cited by
-
Molecular Cloning and Expression Analysis of the Cryptochrome Gene CiPlant-CRY1 in Antarctic Ice Alga Chlamydomonas sp. ICE-L.Plants (Basel). 2022 Aug 26;11(17):2213. doi: 10.3390/plants11172213. Plants (Basel). 2022. PMID: 36079595 Free PMC article.
-
Structure of the bifunctional cryptochrome aCRY from Chlamydomonas reinhardtii.Nucleic Acids Res. 2018 Sep 6;46(15):8010-8022. doi: 10.1093/nar/gky621. Nucleic Acids Res. 2018. PMID: 30032195 Free PMC article.
-
Mechanisms of Cryptochrome-Mediated Photoresponses in Plants.Annu Rev Plant Biol. 2020 Apr 29;71:103-129. doi: 10.1146/annurev-arplant-050718-100300. Epub 2020 Mar 13. Annu Rev Plant Biol. 2020. PMID: 32169020 Free PMC article.
-
Algal photoreceptors: in vivo functions and potential applications.Planta. 2014 Jan;239(1):1-26. doi: 10.1007/s00425-013-1962-5. Epub 2013 Oct 1. Planta. 2014. PMID: 24081482 Review.
-
Dynamics and mechanism of DNA repair by a bifunctional cryptochrome.Proc Natl Acad Sci U S A. 2024 Dec 10;121(50):e2417633121. doi: 10.1073/pnas.2417633121. Epub 2024 Dec 2. Proc Natl Acad Sci U S A. 2024. PMID: 39621923 Free PMC article.
References
-
- Balland V., Byrdin M., Eker A.P., Ahmad M., Brettel K. (2009). What makes the difference between a cryptochrome and DNA photolyase? A spectroelectrochemical comparison of the flavin redox transitions. J. Am. Chem. Soc. 131: 426–427 - PubMed
-
- Banerjee R., Schleicher E., Meier S., Viana R.M., Pokorny R., Ahmad M., Bittl R., Batschauer A. (2007). The signaling state of Arabidopsis cryptochrome 2 contains flavin semiquinone. J. Biol. Chem. 282: 14916–14922 - PubMed
-
- Berndt A., Kottke T., Breitkreuz H., Dvorsky R., Hennig S., Alexander M., Wolf E. (2007). A novel photoreaction mechanism for the circadian blue light photoreceptor Drosophila cryptochrome. J. Biol. Chem. 282: 13011–13021 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
