

Microtubule-targeting drugs rescue axonal swellings in cortical neurons from spastin knockout mice
- PMID: 22773755
- PMCID: PMC3529340
- DOI: 10.1242/dmm.008946
Microtubule-targeting drugs rescue axonal swellings in cortical neurons from spastin knockout mice
Abstract
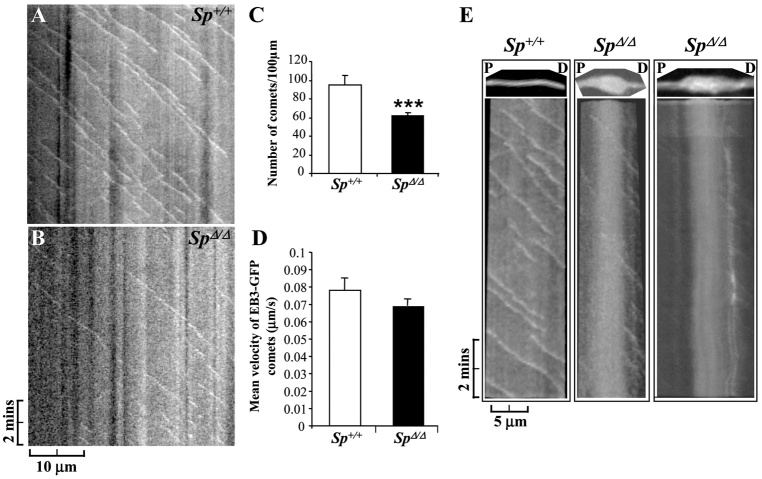
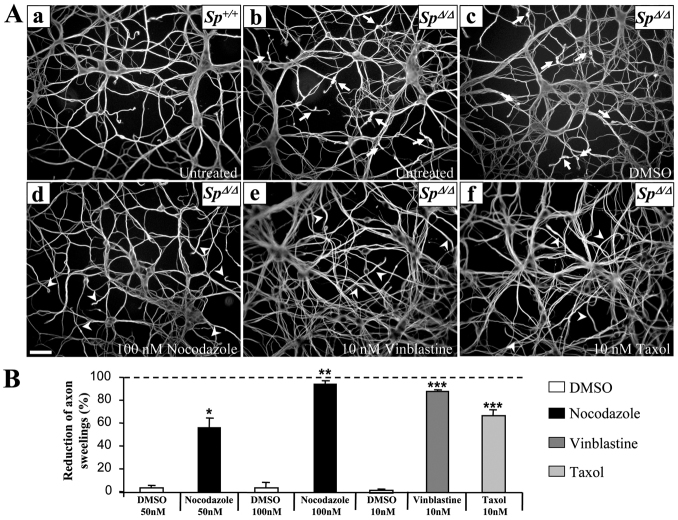
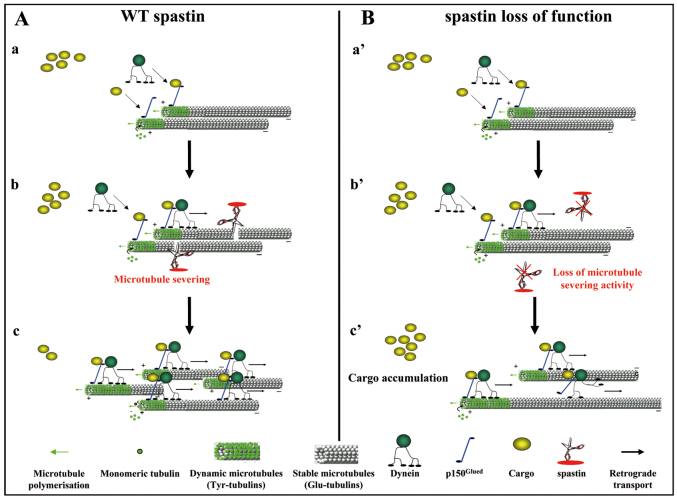
Mutations in SPG4, encoding the microtubule-severing protein spastin, are responsible for the most frequent form of hereditary spastic paraplegia (HSP), a heterogeneous group of genetic diseases characterized by degeneration of the corticospinal tracts. We previously reported that mice harboring a deletion in Spg4, generating a premature stop codon, develop progressive axonal degeneration characterized by focal axonal swellings associated with impaired axonal transport. To further characterize the molecular and cellular mechanisms underlying this mutant phenotype, we have assessed microtubule dynamics and axonal transport in primary cultures of cortical neurons from spastin-mutant mice. We show an early and marked impairment of microtubule dynamics all along the axons of spastin-deficient cortical neurons, which is likely to be responsible for the occurrence of axonal swellings and cargo stalling. Our analysis also reveals that a modulation of microtubule dynamics by microtubule-targeting drugs rescues the mutant phenotype of cortical neurons. Together, these results contribute to a better understanding of the pathogenesis of SPG4-linked HSP and ascertain the influence of microtubule-targeted drugs on the early axonal phenotype in a mouse model of the disease.
Figures
Similar articles
-
Pathogenic mutation of spastin has gain-of-function effects on microtubule dynamics.J Neurosci. 2014 Jan 29;34(5):1856-67. doi: 10.1523/JNEUROSCI.3309-13.2014. J Neurosci. 2014. PMID: 24478365 Free PMC article.
-
Loss of spastin function results in disease-specific axonal defects in human pluripotent stem cell-based models of hereditary spastic paraplegia.Stem Cells. 2014 Feb;32(2):414-23. doi: 10.1002/stem.1569. Stem Cells. 2014. PMID: 24123785 Free PMC article.
-
Direct evidence for axonal transport defects in a novel mouse model of mutant spastin-induced hereditary spastic paraplegia (HSP) and human HSP patients.J Neurochem. 2009 Jul;110(1):34-44. doi: 10.1111/j.1471-4159.2009.06104.x. Epub 2009 Apr 22. J Neurochem. 2009. PMID: 19453301
-
Hereditary spastic paraplegia SPG4: what is known and not known about the disease.Brain. 2015 Sep;138(Pt 9):2471-84. doi: 10.1093/brain/awv178. Epub 2015 Jun 20. Brain. 2015. PMID: 26094131 Free PMC article. Review.
-
The AAA ATPase spastin links microtubule severing to membrane modelling.Biochim Biophys Acta. 2012 Jan;1823(1):192-7. doi: 10.1016/j.bbamcr.2011.08.010. Epub 2011 Aug 25. Biochim Biophys Acta. 2012. PMID: 21888932 Review.
Cited by
-
Beyond taxol: microtubule-based treatment of disease and injury of the nervous system.Brain. 2013 Oct;136(Pt 10):2937-51. doi: 10.1093/brain/awt153. Epub 2013 Jun 27. Brain. 2013. PMID: 23811322 Free PMC article. Review.
-
Patient-Derived Stem Cell Models in SPAST HSP: Disease Modelling and Drug Discovery.Brain Sci. 2018 Jul 31;8(8):142. doi: 10.3390/brainsci8080142. Brain Sci. 2018. PMID: 30065201 Free PMC article. Review.
-
FIGNL1 associates with KIF1Bβ and BICD1 to restrict dynein transport velocity during axon navigation.J Cell Biol. 2019 Oct 7;218(10):3290-3306. doi: 10.1083/jcb.201805128. Epub 2019 Sep 19. J Cell Biol. 2019. PMID: 31541015 Free PMC article.
-
Pathogenic mutation of spastin has gain-of-function effects on microtubule dynamics.J Neurosci. 2014 Jan 29;34(5):1856-67. doi: 10.1523/JNEUROSCI.3309-13.2014. J Neurosci. 2014. PMID: 24478365 Free PMC article.
-
Axonal autophagosome maturation defect through failure of ATG9A sorting underpins pathology in AP-4 deficiency syndrome.Autophagy. 2020 Mar;16(3):391-407. doi: 10.1080/15548627.2019.1615302. Epub 2019 May 29. Autophagy. 2020. PMID: 31142229 Free PMC article.
References
-
- Baas P. W., Qiang L. (2005). Neuronal microtubules: when the MAP is the roadblock. Trends Cell Biol. 15, 183–187 - PubMed
-
- Baas P. W., Karabay A., Qiang L. (2005). Microtubules cut and run. Trends Cell Biol. 15, 518–524 - PubMed
-
- Charvin D., Cifuentes-Diaz C., Fonknechten N., Joshi V., Hazan J., Melki J., Betuing S. (2003). Mutations of SPG4 are responsible for a loss of function of spastin, an abundant neuronal protein localized in the nucleus. Hum. Mol. Genet. 12, 71–78 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
