

Does interictal synchronization influence ictogenesis?
- PMID: 22776544
- PMCID: PMC4878915
- DOI: 10.1016/j.neuropharm.2012.06.044
Does interictal synchronization influence ictogenesis?
Abstract
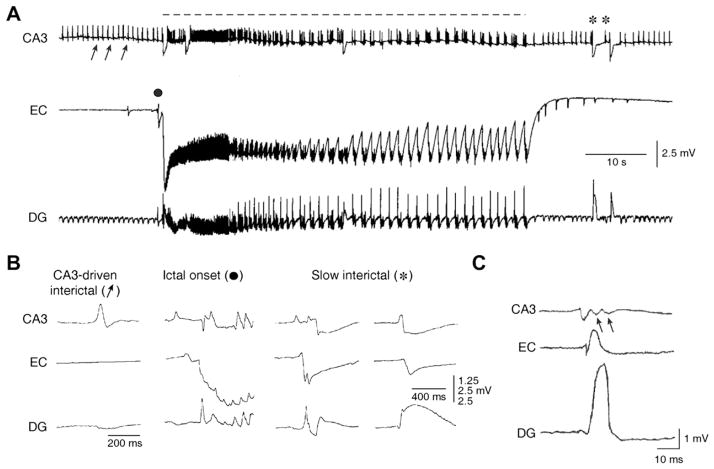
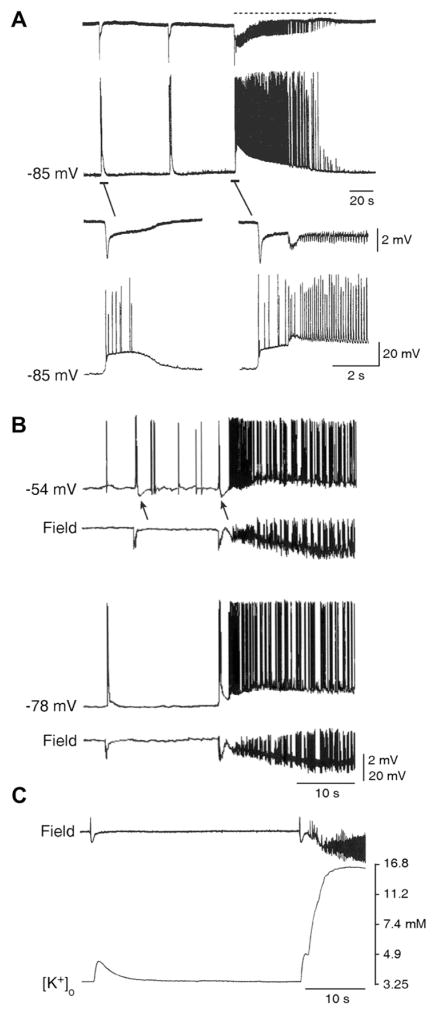
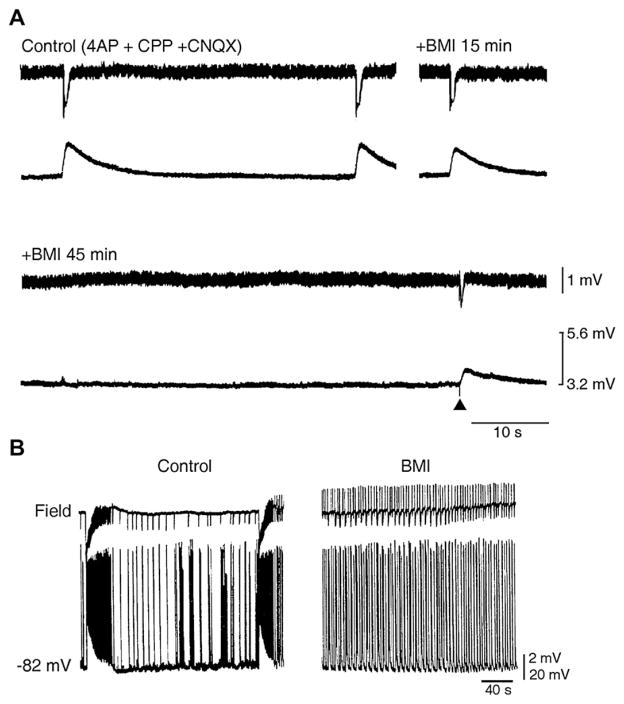
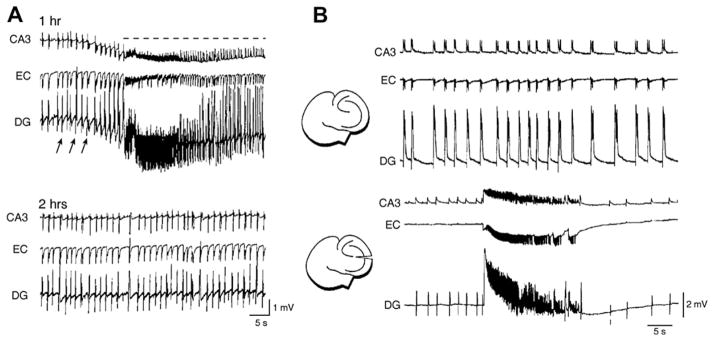
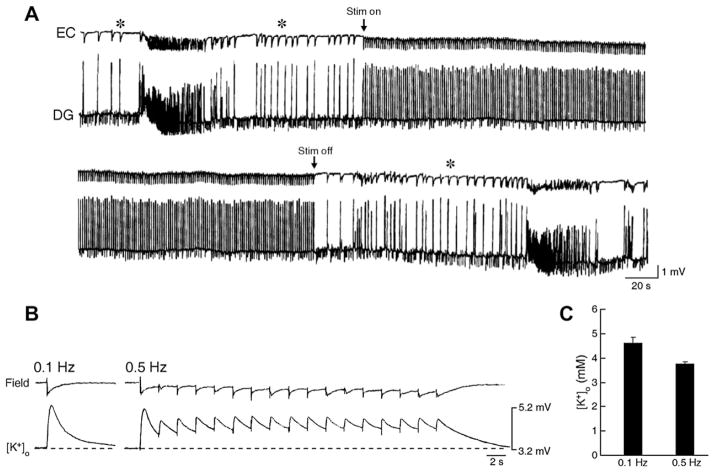
The EEG recorded from epileptic patients presents with interictal discharges that are not associated with detectable clinical symptoms but are valuable for diagnostic purposes. Experimental studies have shown that interictal discharges and ictal events (i.e., seizures) are characterized intracellularly by similar (but for duration) neuronal depolarizations leading to sustained action potential firing, thus indicating that they may share similar cellular and pharmacological mechanisms. It has also been proposed that interictal discharges may herald the onset of electrographic seizures, but other studies have demonstrated that interictal events interfere with the occurrence of ictal activity. The relationship between interictal and ictal activity thus remains ambiguous. Here we will review this issue in animal models of limbic seizures that are electrographically close to those seen in TLE patients. In particular we will: (i) focus on the electrophysiological and pharmacological characteristics of, at least, two types of interictal discharge; (ii) propose that they play opposite roles in leading to ictogenesis; and (iii) discuss the possibility that mimicking one of these two types of interictal activity by low frequency repetitive stimulation can control ictogenesis. Finally, we will also review evidence indicating that specific types of interictal discharge may play a role in epileptogenesis. This article is part of the Special Issue entitled 'New Targets and Approaches to the Treatment of Epilepsy'.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Ayala GF, Dichter M, Gumnit RJ, Matsumoto H, Spencer WA. Genesis of epileptic interictal spikes. New knowledge of cortical feedback systems suggests a neurophysiological explanation of brief paroxysms. Brain Res. 1973;1973(52):1–17. - PubMed
-
- Barbarosie M, Louvel J, D’Antuono M, Kurcewicz I, Avoli M. Masking synchronous GABA-mediated potentials controls limbic seizures. Epilepsia. 2002;43:1469–1479. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
