

Ca(2+) influx and neurotransmitter release at ribbon synapses
- PMID: 22776680
- PMCID: PMC3468149
- DOI: 10.1016/j.ceca.2012.06.004
Ca(2+) influx and neurotransmitter release at ribbon synapses
Abstract
Ca(2+) influx through voltage-gated Ca(2+) channels triggers the release of neurotransmitters at presynaptic terminals. Some sensory receptor cells in the peripheral auditory and visual systems have specialized synapses that express an electron-dense organelle called a synaptic ribbon. Like conventional synapses, ribbon synapses exhibit SNARE-mediated exocytosis, clathrin-mediated endocytosis, and short-term plasticity. However, unlike non-ribbon synapses, voltage-gated L-type Ca(2+) channel opening at ribbon synapses triggers a form of multiquantal release that can be highly synchronous. Furthermore, ribbon synapses appear to be specialized for fast and high throughput exocytosis controlled by graded membrane potential changes. Here we will discuss some of the basic aspects of synaptic transmission at different types of ribbon synapses, and we will emphasize recent evidence that auditory and retinal ribbon synapses have marked differences. This will lead us to suggest that ribbon synapses are specialized for particular operating ranges and frequencies of stimulation. We propose that different types of ribbon synapses transfer diverse rates of sensory information by expressing a particular repertoire of critical components, and by placing them at precise and strategic locations, so that a continuous supply of primed vesicles and Ca(2+) influx leads to fast, accurate, and ongoing exocytosis.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
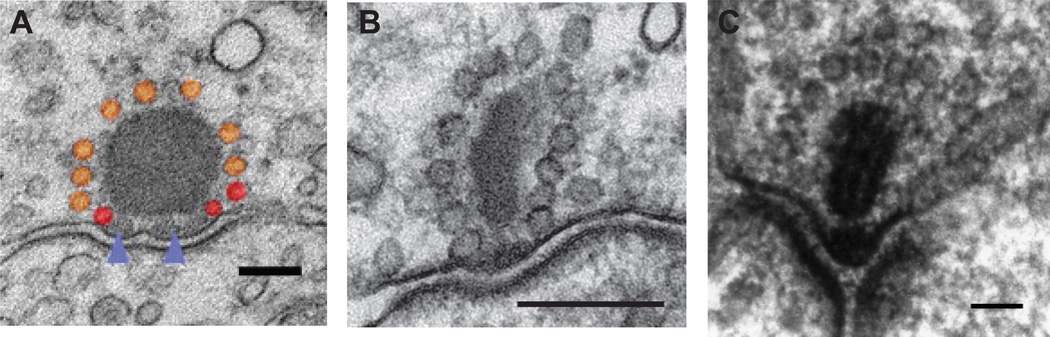
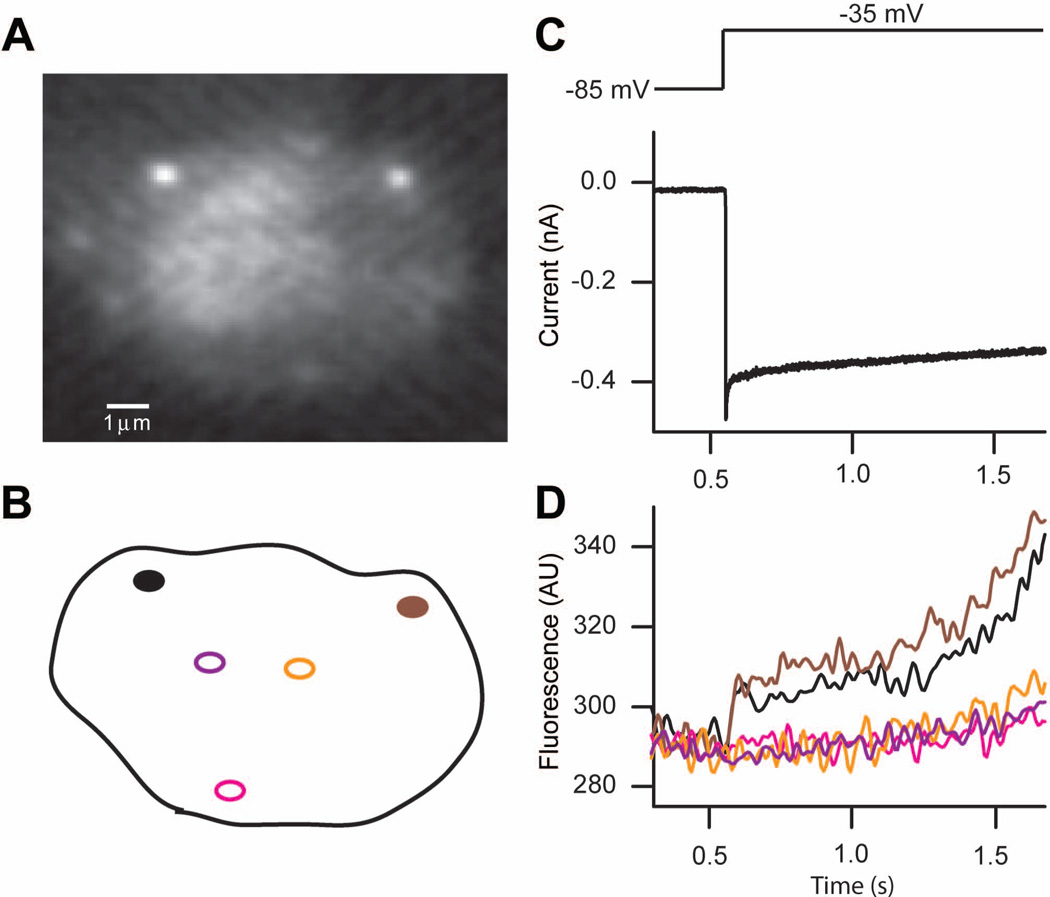

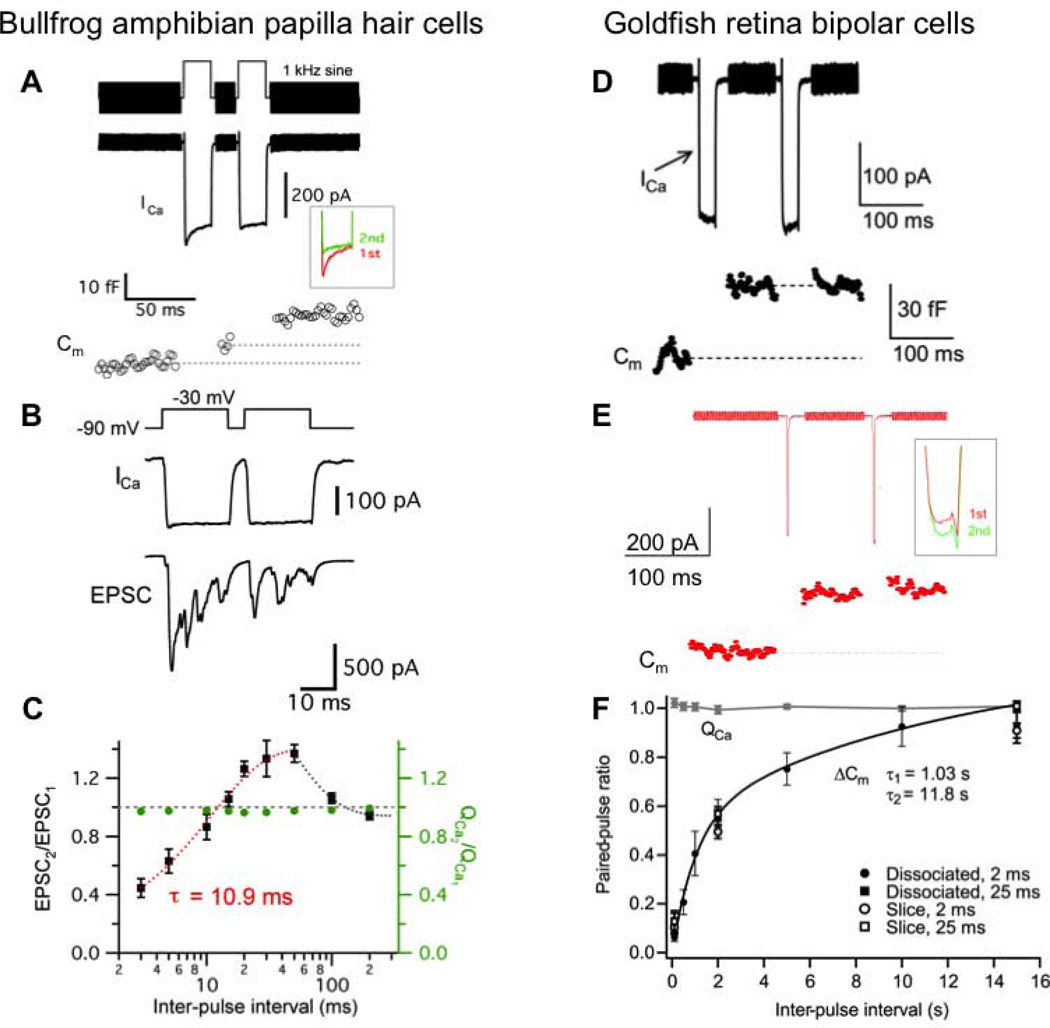
Similar articles
-
Visualizing synaptic vesicle turnover and pool refilling driven by calcium nanodomains at presynaptic active zones of ribbon synapses.Proc Natl Acad Sci U S A. 2014 Jun 10;111(23):8655-60. doi: 10.1073/pnas.1323962111. Epub 2014 May 27. Proc Natl Acad Sci U S A. 2014. PMID: 24912160 Free PMC article.
-
The molecular architecture of ribbon presynaptic terminals.Mol Neurobiol. 2009 Apr;39(2):130-48. doi: 10.1007/s12035-009-8058-z. Epub 2009 Mar 3. Mol Neurobiol. 2009. PMID: 19253034 Free PMC article. Review.
-
Sharp Ca²⁺ nanodomains beneath the ribbon promote highly synchronous multivesicular release at hair cell synapses.J Neurosci. 2011 Nov 16;31(46):16637-50. doi: 10.1523/JNEUROSCI.1866-11.2011. J Neurosci. 2011. PMID: 22090491 Free PMC article.
-
Neurotransmitter release at ribbon synapses in the retina.Immunol Cell Biol. 2000 Aug;78(4):442-6. doi: 10.1046/j.1440-1711.2000.00923.x. Immunol Cell Biol. 2000. PMID: 10947871 Review.
-
Balancing presynaptic release and endocytic membrane retrieval at hair cell ribbon synapses.FEBS Lett. 2018 Nov;592(21):3633-3650. doi: 10.1002/1873-3468.13258. Epub 2018 Oct 9. FEBS Lett. 2018. PMID: 30251250 Review.
Cited by
-
Autonomous push button-controlled rapid insulin release from a piezoelectrically activated subcutaneous cell implant.Sci Adv. 2022 Jun 17;8(24):eabm4389. doi: 10.1126/sciadv.abm4389. Epub 2022 Jun 15. Sci Adv. 2022. PMID: 35704573 Free PMC article.
-
Phase-Locking Requires Efficient Ca2+ Extrusion at the Auditory Hair Cell Ribbon Synapse.J Neurosci. 2021 Feb 24;41(8):1625-1635. doi: 10.1523/JNEUROSCI.1324-18.2020. Epub 2021 Jan 14. J Neurosci. 2021. PMID: 33446517 Free PMC article.
-
Synaptic ribbons foster active zone stability and illumination-dependent active zone enrichment of RIM2 and Cav1.4 in photoreceptor synapses.Sci Rep. 2020 Apr 6;10(1):5957. doi: 10.1038/s41598-020-62734-0. Sci Rep. 2020. PMID: 32249787 Free PMC article.
-
Bridging the gap between presynaptic hair cell function and neural sound encoding.Elife. 2024 Dec 24;12:RP93749. doi: 10.7554/eLife.93749. Elife. 2024. PMID: 39718472 Free PMC article.
-
Harmonin enhances voltage-dependent facilitation of Cav1.3 channels and synchronous exocytosis in mouse inner hair cells.J Physiol. 2013 Jul 1;591(13):3253-69. doi: 10.1113/jphysiol.2013.254367. Epub 2013 Apr 22. J Physiol. 2013. PMID: 23613530 Free PMC article.
References
-
- Lenzi D, von Gersdorff H. Structure suggests function: the case for synaptic ribbons as exocytotic nanomachines. Bioessays. 2001;23:831–840. - PubMed
-
- Liberman MC. Morphological differences among radial afferent fibers in the cat cochlea: an electron-microscopic study of serial sections. Hear Res. 1980;3:45–63. - PubMed
-
- von Gersdorff H. Synaptic ribbons: versatile signal transducers. Neuron. 2001;29:7–10. - PubMed
-
- Parsons TD, Sterling P. Synaptic ribbon: Conveyor belt or safety belt? Neuron. 2003;37:379–382. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
