

Cannabinoid receptor activation correlates with the proapoptotic action of the β2-adrenergic agonist (R,R')-4-methoxy-1-naphthylfenoterol in HepG2 hepatocarcinoma cells
- PMID: 22776956
- PMCID: PMC3464034
- DOI: 10.1124/jpet.112.195206
Cannabinoid receptor activation correlates with the proapoptotic action of the β2-adrenergic agonist (R,R')-4-methoxy-1-naphthylfenoterol in HepG2 hepatocarcinoma cells
Abstract
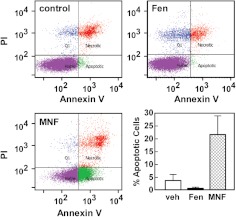
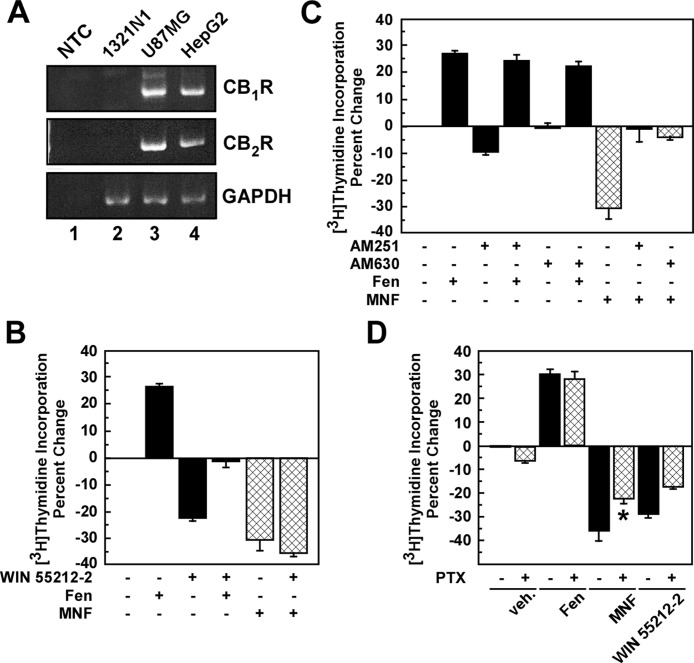
Inhibition of cell proliferation by fenoterol and fenoterol derivatives in 1321N1 astrocytoma cells is consistent with β(2)-adrenergic receptor (β(2)-AR) stimulation. However, the events that result in fenoterol-mediated control of cell proliferation in other cell types are not clear. Here, we compare the effect of the β(2)-AR agonists (R,R')-fenoterol (Fen) and (R,R')-4-methoxy-1-naphthylfenoterol (MNF) on signaling and cell proliferation in HepG2 hepatocarcinoma cells by using Western blotting and [(3)H]thymidine incorporation assays. Despite the expression of β(2)-AR, no cAMP accumulation was observed when cells were stimulated with isoproterenol or Fen, although the treatment elicited both mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt activation. Unexpectedly, isoproterenol and Fen promoted HepG2 cell growth, but MNF reduced proliferation together with increased apoptosis. The mitogenic responses of Fen were attenuated by 3-(isopropylamino)-1-[(7-methyl-4-indanyl)oxy]butan-2-ol (ICI 118,551), a β(2)-AR antagonist, whereas those of MNF were unaffected. Because of the coexpression of β(2)-AR and cannabinoid receptors (CBRs) and their impact on HepG2 cell proliferation, these Gα(i)/Gα(o)-linked receptors may be implicated in MNF signaling. Cell treatment with (R)-(+)-[2,3-dihydro-5-methyl-3-(4-morpholinylmethyl)pyrrolo[1,2,3-de]-1,4-benzoxazin-6-yl]-1-napthalenylmethanone (WIN 55,212-2), a synthetic agonist of CB(1)R and CB(2)R, led to growth inhibition, whereas inverse agonists of these receptors blocked MNF mitogenic responses without affecting Fen signaling. MNF responses were sensitive to pertussis toxin. The β(2)-AR-deficient U87MG cells were refractory to Fen, but responsive to the antiproliferative actions of MNF and WIN 55,212-2. The data indicate that the presence of the naphthyl moiety in MNF results in functional coupling to the CBR pathway, providing one of the first examples of a dually acting β(2)-AR-CBR ligand.
Figures
References
-
- Aguado T, Carracedo A, Julien B, Velasco G, Milman G, Mechoulam R, Alvarez L, Guzmán M, Galve-Roperh I. (2007) Cannabinoids induce glioma stem-like cell differentiation and inhibit gliomagenesis. J Biol Chem 282:6854–6862 - PubMed
-
- Ahn S, Maudsley S, Luttrell LM, Lefkowitz RJ, Daaka Y. (1999) Src-mediated tyrosine phosphorylation of dynamin is required for β2-adrenergic receptor internalization and mitogen- activated protein kinase signaling. J Biol Chem 274:1185–1188 - PubMed
-
- Audet M, Bouvier M. (2008) Insights into signaling from the β2-adrenergic receptor structure. Nat Chem Biol 4:397–403 - PubMed
-
- Biswas KK, Sarker KP, Abeyama K, Kawahara K, Iino S, Otsubo Y, Saigo K, Izumi H, Hashiguchi T, Yamakuchi M, et al. (2003) Membrane cholesterol but not putative receptors mediates anandamide-induced hepatocyte apoptosis. Hepatology 38:1167–1177 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
