

High-temperature requirement protein A4 (HtrA4) suppresses the fusogenic activity of syncytin-1 and promotes trophoblast invasion
- PMID: 22778138
- PMCID: PMC3430202
- DOI: 10.1128/MCB.00223-12
High-temperature requirement protein A4 (HtrA4) suppresses the fusogenic activity of syncytin-1 and promotes trophoblast invasion
Abstract
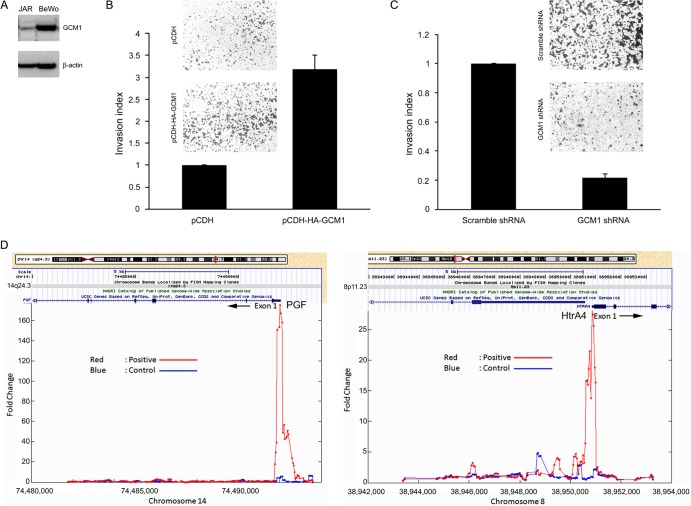
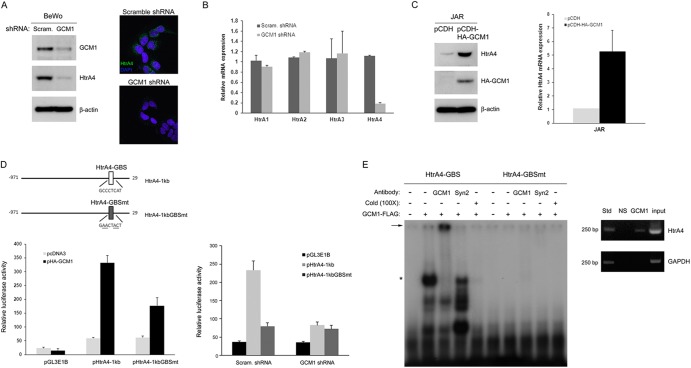
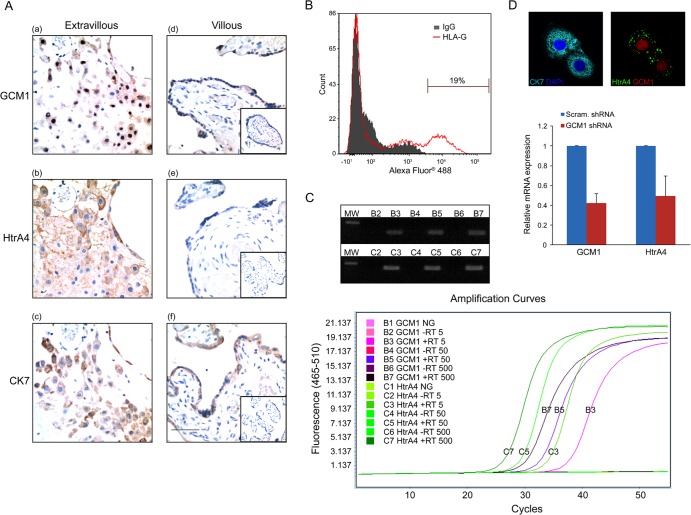
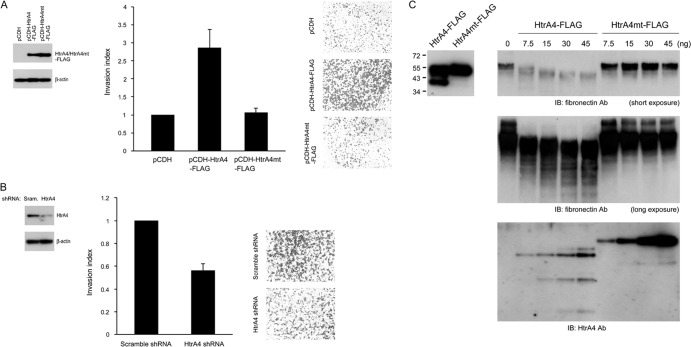
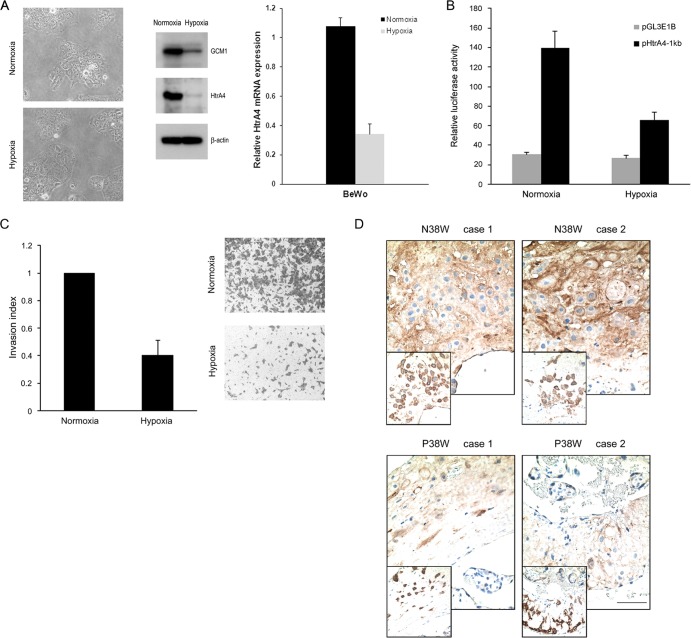
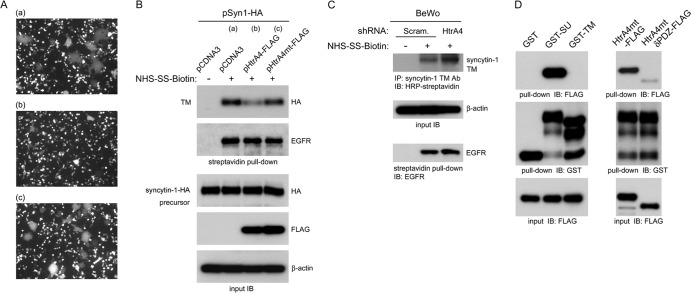
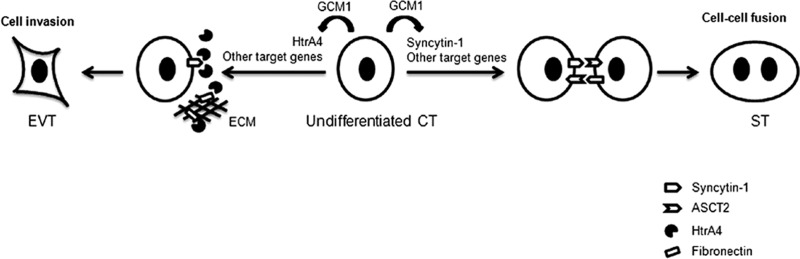
Cell-cell fusion and cell invasion are essential for placental development. Human cytotrophoblasts in the chorionic villi may undergo cell-cell fusion to form syncytiotrophoblasts to facilitate nutrient-gas exchange or differentiate into extravillous trophoblasts (EVTs) to facilitate maternal-fetal circulation. The placental transcription factor glial cells missing 1 (GCM1) regulates syncytin-1 and -2 expression to mediate trophoblast fusion. Interestingly, GCM1 and syncytin-1 are also expressed in EVTs with unknown physiological functions. In this study, we performed chromatin immunoprecipitation-on-chip (ChIP-chip) analysis and identified the gene for high-temperature requirement protein A4 (HtrA4) as a GCM1 target gene, which encodes a serine protease facilitating cleavage of fibronectin and invasion of placental cells. Importantly, HtrA4 is immunolocalized in EVTs at the maternal-fetal interface, and its expression is decreased by hypoxia and in preeclampsia, a pregnancy complication associated with placental hypoxia and shallow trophoblast invasion. We further demonstrate that HtrA4 interacts with syncytin-1 and suppresses cell-cell fusion. Therefore, HtrA4 may be crucial for EVT differentiation by playing a dual role in prevention of cell-cell fusion of EVTs and promotion of their invasion into the uterus. Our study reveals a novel function of GCM1 and HtrA4 in regulation of trophoblast invasion and that abnormal HrtA4 expression may contribute to shallow trophoblast invasion in preeclampsia.
Figures
References
-
- Ajayi F, et al. 2008. Elevated expression of serine protease HtrA1 in preeclampsia and its role in trophoblast cell migration and invasion. Am. J. Obstet. Gynecol. 199:557.e1–557.e10 - PubMed
-
- Anson-Cartwright L, et al. 2000. The glial cells missing-1 protein is essential for branching morphogenesis in the chorioallantoic placenta. Nat. Genet. 25:311–314 - PubMed
-
- Baczyk D, et al. 2004. Complex patterns of GCM1 mRNA and protein in villous and extravillous trophoblast cells of the human placenta. Placenta 25:553–559 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases