

The autophagy-related protein kinase Atg1 interacts with the ubiquitin-like protein Atg8 via the Atg8 family interacting motif to facilitate autophagosome formation
- PMID: 22778255
- PMCID: PMC3436563
- DOI: 10.1074/jbc.C112.387514
The autophagy-related protein kinase Atg1 interacts with the ubiquitin-like protein Atg8 via the Atg8 family interacting motif to facilitate autophagosome formation
Abstract
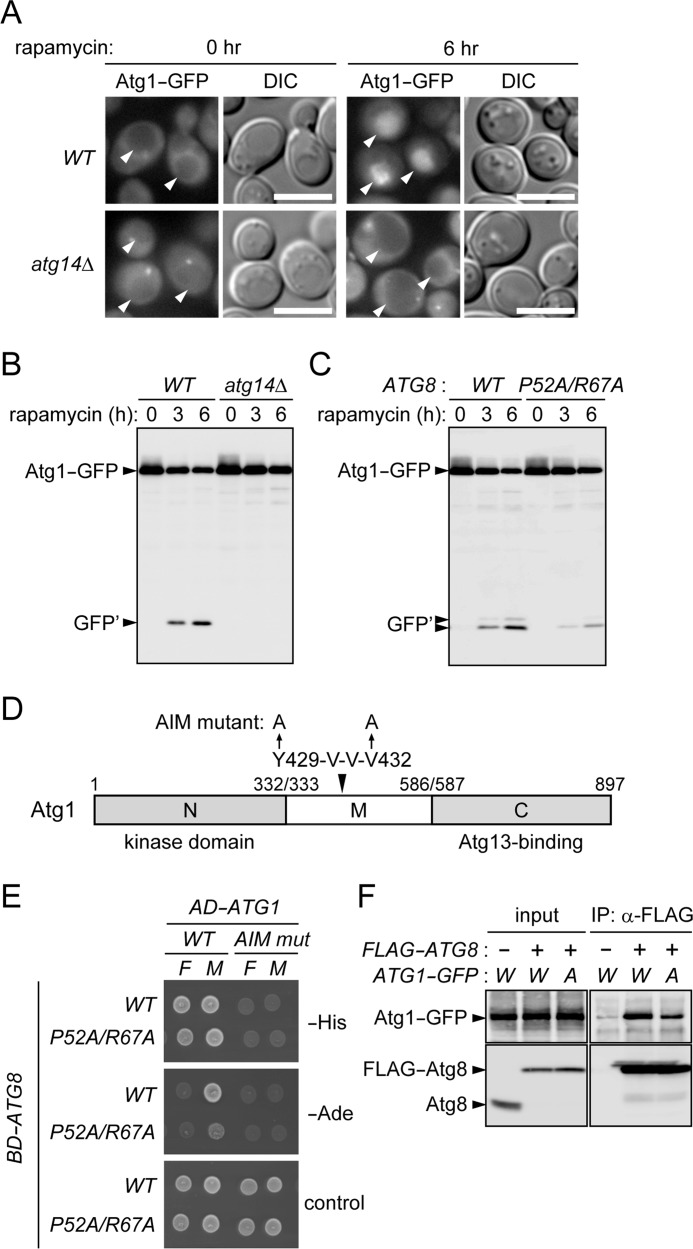
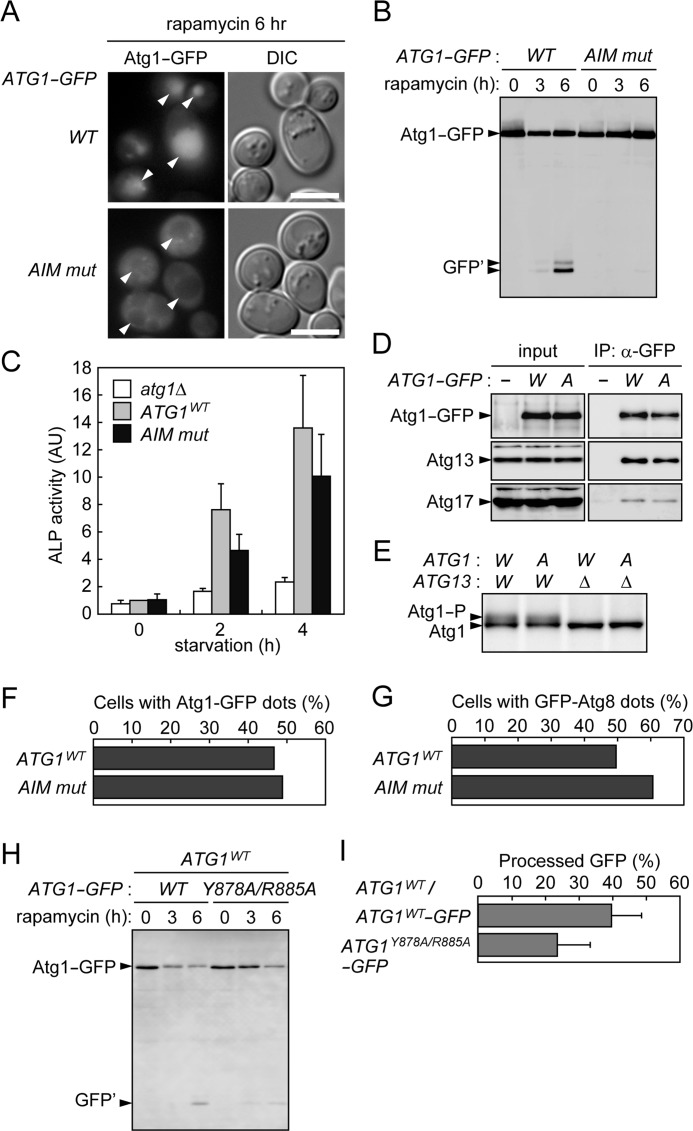
In autophagy, a cup-shaped membrane called the isolation membrane is formed, expanded, and sealed to complete a double membrane-bound vesicle called the autophagosome that encapsulates cellular constituents to be transported to and degraded in the lysosome/vacuole. The formation of the autophagosome requires autophagy-related (Atg) proteins. Atg8 is a ubiquitin-like protein that localizes to the isolation membrane; a subpopulation of this protein remains inside the autophagosome and is transported to the lysosome/vacuole. In the budding yeast Saccharomyces cerevisiae, Atg1 is a serine/threonine kinase that functions in the initial step of autophagosome formation and is also efficiently transported to the vacuole via autophagy. Here, we explore the mechanism and significance of this autophagic transport of Atg1. In selective types of autophagy, receptor proteins recognize degradation targets and also interact with Atg8, via the Atg8 family interacting motif (AIM), to link the targets to the isolation membrane. We find that Atg1 contains an AIM and directly interacts with Atg8. Mutations in the AIM disrupt this interaction and abolish vacuolar transport of Atg1. These results suggest that Atg1 associates with the isolation membrane by binding to Atg8, resulting in its incorporation into the autophagosome. We also show that mutations in the Atg1 AIM cause a significant defect in autophagy, without affecting the functions of Atg1 implicated in triggering autophagosome formation. We propose that in addition to its essential function in the initial stage, Atg1 also associates with the isolation membrane to promote its maturation into the autophagosome.
Figures
References
-
- Nakatogawa H., Suzuki K., Kamada Y., Ohsumi Y. (2009) Dynamics and diversity in autophagy mechanisms: lessons from yeast. Nat. Rev. Mol. Cell Biol. 10, 458–467 - PubMed
-
- Weidberg H., Shvets E., Elazar Z. (2011) Biogenesis and cargo selectivity of autophagosomes. Annu. Rev. Biochem. 80, 125–156 - PubMed
-
- Ichimura Y., Kirisako T., Takao T., Satomi Y., Shimonishi Y., Ishihara N., Mizushima N., Tanida I., Kominami E., Ohsumi M., Noda T., Ohsumi Y. (2000) A ubiquitin-like system mediates protein lipidation. Nature 408, 488–492 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
