

Review
doi: 10.1083/jcb.201112044.
The cell biology of disease: Acute promyelocytic leukemia, arsenic, and PML bodies
Affiliations
- PMID: 22778276
- PMCID: PMC3392943
- DOI: 10.1083/jcb.201112044
Item in Clipboard
Review
The cell biology of disease: Acute promyelocytic leukemia, arsenic, and PML bodies
J Cell Biol.
.
Abstract
Acute promyelocytic leukemia (APL) is driven by a chromosomal translocation whose product, the PML/retinoic acid (RA) receptor α (RARA) fusion protein, affects both nuclear receptor signaling and PML body assembly. Dissection of APL pathogenesis has led to the rediscovery of PML bodies and revealed their role in cell senescence, disease pathogenesis, and responsiveness to treatment. APL is remarkable because of the fortuitous identification of two clinically effective therapies, RA and arsenic, both of which degrade PML/RARA oncoprotein and, together, cure APL. Analysis of arsenic-induced PML or PML/RARA degradation has implicated oxidative stress in the biogenesis of nuclear bodies and SUMO in their degradation.
Figures
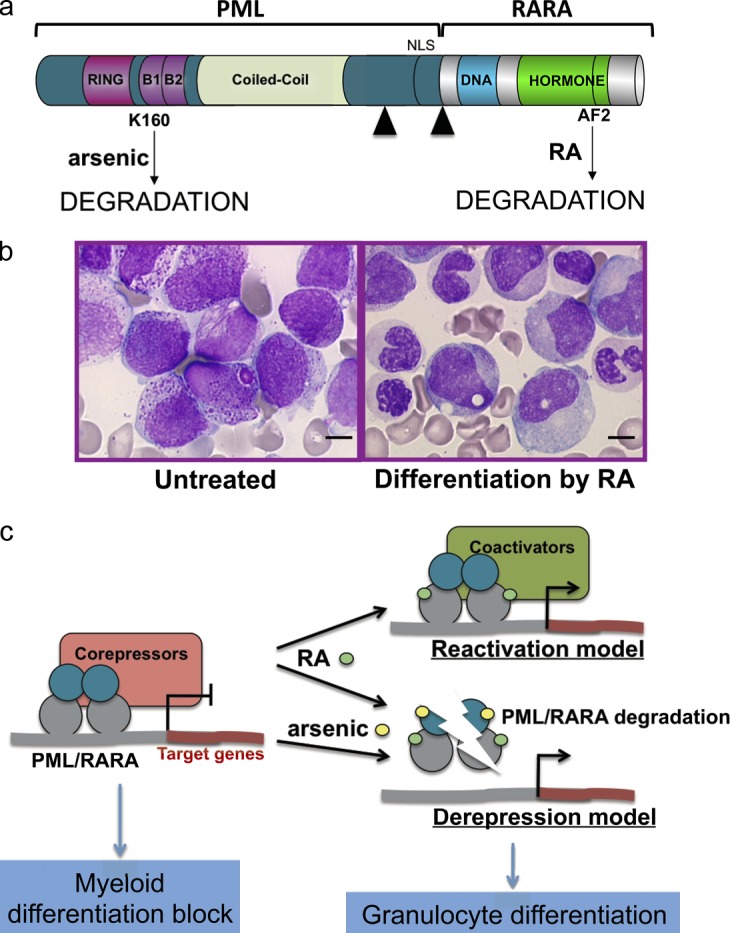
PML/RARA, RA, arsenic, and APL. (a) The PML/RARA fusion protein with the main functional domains of PML and RARA. Note that RA targets the RARA moiety, and arsenic targets the PML moiety, both leading to PML/RARA degradation. Black arrowheads denote the different fusion points. DNA, DNA-binding domain; AF2, activating function 2. (b) RA treatment in vivo elicits the differentiation of leukemia cells (reproduced from Zhu et al. [2002] with permission from Nature Publishing Group). Bars, 5 µm. (c) Current models of RA or arsenic action on transcriptional control and differentiation through gene activation (top) or derepression via degradation (bottom).
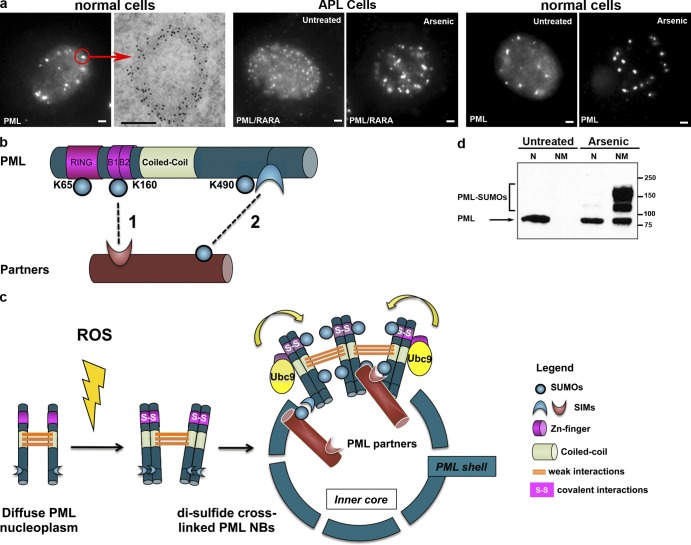
PML and NBs. (a) PML NBs in normal and APL cells. From left to right: (normal cells) immunofluorescence and electron microscopy views in CHO cells stably expressing PML. (left) PML is both diffusely distributed in the nucleoplasm and aggregated in NBs. The red arrow points to an individual body, analyzed by electron microscopy. Bar, 1 µm. (right) anti-PML antibodies, labeled with gold particles, show PML distribution in the electron-dense NB shell. Bar, 0.5 µm. (APL cells) NB disruption into micropunctuations by PML/RARA expression (left) and reformation upon arsenic treatment (right). (normal cells) Arsenic controls NB aggregation in non-APL cells. CHO cells stably expressing PML were treated (right) or not treated (left) with 1 µM As2O3 for 1 h. Note the disappearance of the diffuse nuclear staining and increased size of NBs. (b) PML structure and SUMO-induced modifications. Interactions of partner proteins with SUMO–SIM are shown as dotted lines. PML primarily interacts with its partners through K160 sumoylation (1); NB association of sumoylated partners is secured by subsequent SIM–SUMO reciprocal interactions between partners and possibly PML SIM (2). (c) PML cross-linking by disulfide bounds underlies formation of the matrix-associated shell, and polarized SUMO–SIM interactions recruit partner proteins within NBs. PML multimers recruit Ubc9, leading to sumoylation of PML and possibly its partners. S, sulfur. (d) PML targeting to the insoluble nuclear matrix (NM) after arsenic exposure. Reproduced from Lallemand-Breitenbach et al. (2001). Molecular masses are given in kilodaltons. N, nucleoplasm.
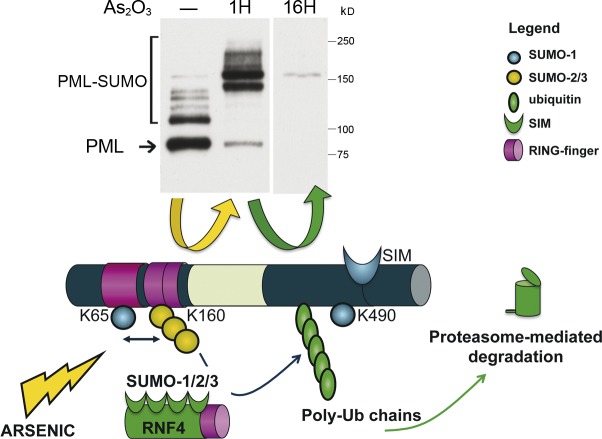
PML or PML/RARA degradation by arsenic trioxide. Arsenic enhances multi- and/or polysumoylation by SUMO1/2/3 on K65 and K160, but not K490, as demonstrated by Western blotting after short treatment of CHO cells stably expressing PML (top, middle lane); PML proteins are degraded after longer exposure to arsenic (top, right lane). The ubiquitin E3 ligase RFN4 comprises four SIMs and is thus recruited onto dimerized, mesh-associated, hypersumoylated PML to induce its polyubiquitination (poly-Ub). PML is finally degraded by the proteasome machinery, which is also recruited onto NBs. The yellow arrow depicts the events occurring in the first hour after arsenic, and the green one shows subsequent degradation (corresponding to polyubiquitination).
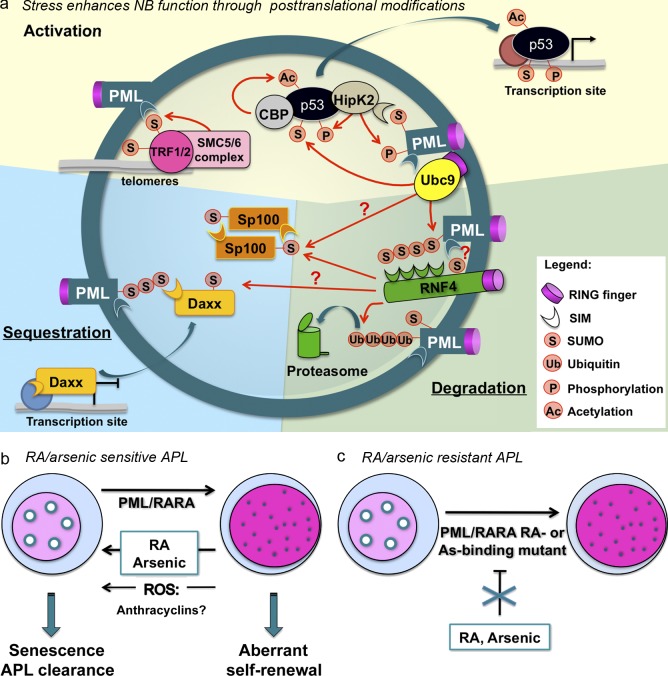
PML NB partners and NB-triggered biological responses. (a) PML oxidation elicits PML NB formation. Enzymes, such as SUMO E2 Ubc9, SUMO E3 SMC5/6 subunits, ubiquitin E3 RNF4, the acetyltransferase CBP, or the kinase HIPK2, are recruited onto NBs. Concentrating enzymes and their substrates supports posttranslational modifications of partner proteins, such as SP100, the structural maintenance or chromosome (SMC) complex involved in alternative lengthening telomeres, DAXX, or p53. These modifications affect activity and stability of PML partners and/or lead to their sequestration within NBs, converging into quiescence (stem cells) or a senescence program. (b) Proposed links between PML NB reformation and APL response to treatment. PML/RARA disrupts NBs, which could be reflected in aberrant self-renewal. RA or arsenic, by degrading PML/RARA, allows reformation of PML NBs, as do reactive oxygen species (ROS) and possibly anthracyclins. (c) In therapy-resistant APLs, as a result of mutations in PML/RARA, therapies fail to restore NBs.
References
-
- Ablain J., Nasr R., Bazarbachi A., de Thé H. 2011. The drug-induced degradation of oncoproteins: an unexpected achilles’ heel of cancer cells? Cancer Discov. 1:117–127 10.1158/2159-8290.CD-11-0087 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
