

Circadian clock protein cryptochrome regulates the expression of proinflammatory cytokines
- PMID: 22778400
- PMCID: PMC3411996
- DOI: 10.1073/pnas.1209965109
Circadian clock protein cryptochrome regulates the expression of proinflammatory cytokines
Abstract
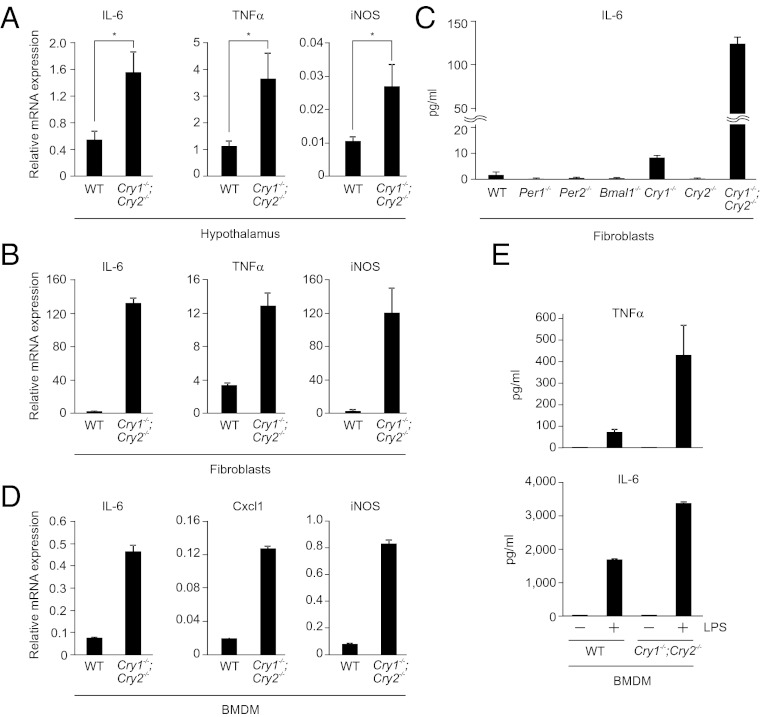
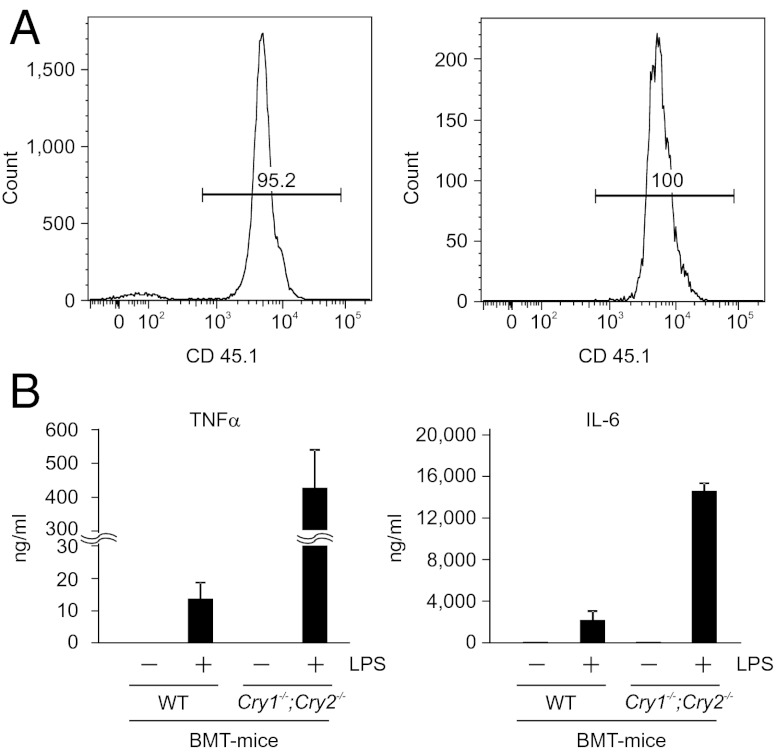
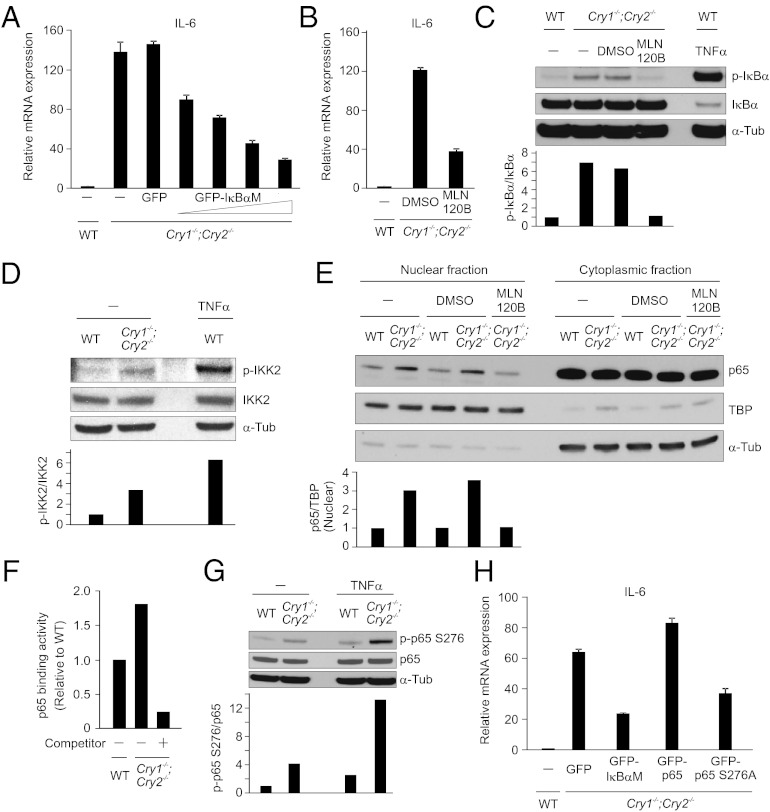
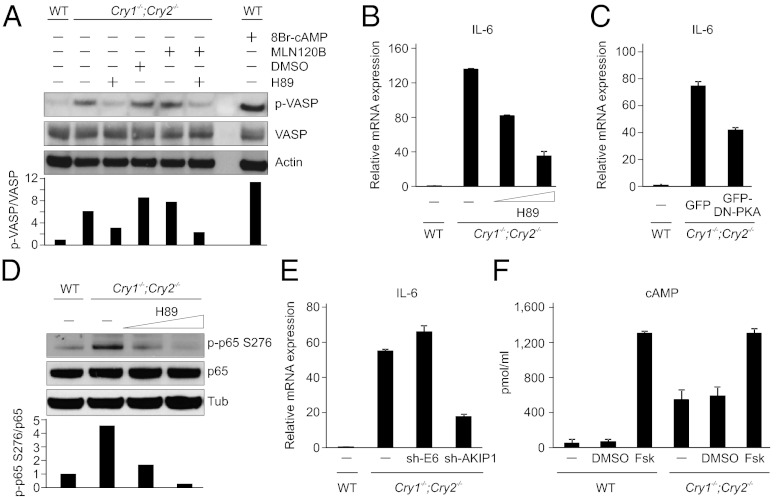
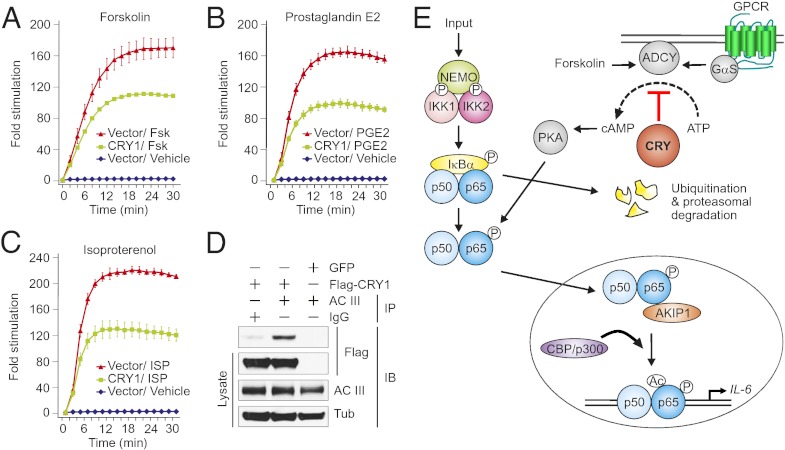
Chronic sleep deprivation perturbs the circadian clock and increases susceptibility to diseases such as diabetes, obesity, and cancer. Increased inflammation is one of the common underlying mechanisms of these diseases, thus raising a hypothesis that circadian-oscillator components may regulate immune response. Here we show that absence of the core clock component protein cryptochrome (CRY) leads to constitutive elevation of proinflammatory cytokines in a cell-autonomous manner. We observed a constitutive NF-κB and protein kinase A (PKA) signaling activation in Cry1(-/-);Cry2(-/-) cells. We further demonstrate that increased phosphorylation of p65 at S276 residue in Cry1(-/-);Cry2(-/-) cells is due to increased PKA signaling activity, likely induced by a significantly high basal level of cAMP, which we detected in these cells. In addition, we report that CRY1 binds to adenylyl cyclase and limits cAMP production. Based on these data, we propose that absence of CRY protein(s) might release its (their) inhibition on cAMP production, resulting in elevated cAMP and increased PKA activation, subsequently leading to NF-κB activation through phosphorylation of p65 at S276. These results offer a mechanistic framework for understanding the link between circadian rhythm disruption and increased susceptibility to chronic inflammatory diseases.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Liu AC, Lewis WG, Kay SA. Mammalian circadian signaling networks and therapeutic targets. Nat Chem Biol. 2007;3:630–639. - PubMed
-
- Saini C, Suter DM, Liani A, Gos P, Schibler U. The mammalian circadian timing system: Synchronization of peripheral clocks. Cold Spring Harb Symp Quant Biol. 2011;76:39–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
