

Eradication of bacterial persisters with antibiotic-generated hydroxyl radicals
- PMID: 22778419
- PMCID: PMC3409745
- DOI: 10.1073/pnas.1203735109
Eradication of bacterial persisters with antibiotic-generated hydroxyl radicals
Abstract
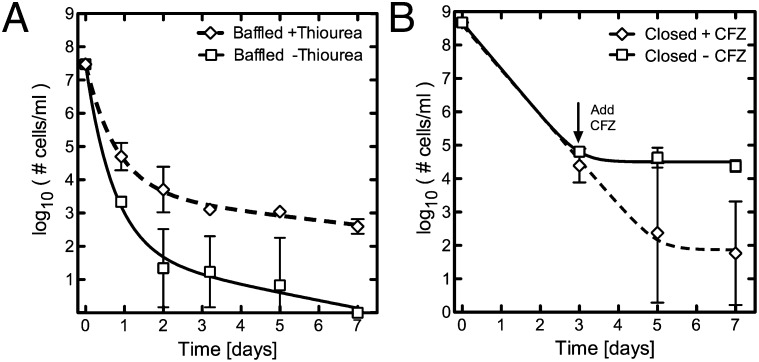
During Mycobacterium tuberculosis infection, a population of bacteria likely becomes refractory to antibiotic killing in the absence of genotypic resistance, making treatment challenging. We describe an in vitro model capable of yielding a phenotypically antibiotic-tolerant subpopulation of cells, often called persisters, within populations of Mycobacterium smegmatis and M. tuberculosis. We find that persisters are distinct from the larger antibiotic-susceptible population, as a small drop in dissolved oxygen (DO) saturation (20%) allows for their survival in the face of bactericidal antibiotics. In contrast, if high levels of DO are maintained, all cells succumb, sterilizing the culture. With increasing evidence that bactericidal antibiotics induce cell death through the production of reactive oxygen species (ROS), we hypothesized that the drop in DO decreases the concentration of ROS, thereby facilitating persister survival, and maintenance of high DO yields sufficient ROS to kill persisters. Consistent with this hypothesis, the hydroxyl-radical scavenger thiourea, when added to M. smegmatis cultures maintained at high DO levels, rescues the persister population. Conversely, the antibiotic clofazimine, which increases ROS via an NADH-dependent redox cycling pathway, successfully eradicates the persister population. Recent work suggests that environmentally induced antibiotic tolerance of bulk populations may result from enhanced antioxidant capabilities. We now show that the small persister subpopulation within a larger antibiotic-susceptible population also shows differential susceptibility to antibiotic-induced hydroxyl radicals. Furthermore, we show that stimulating ROS production can eradicate persisters, thus providing a potential strategy to managing persistent infections.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
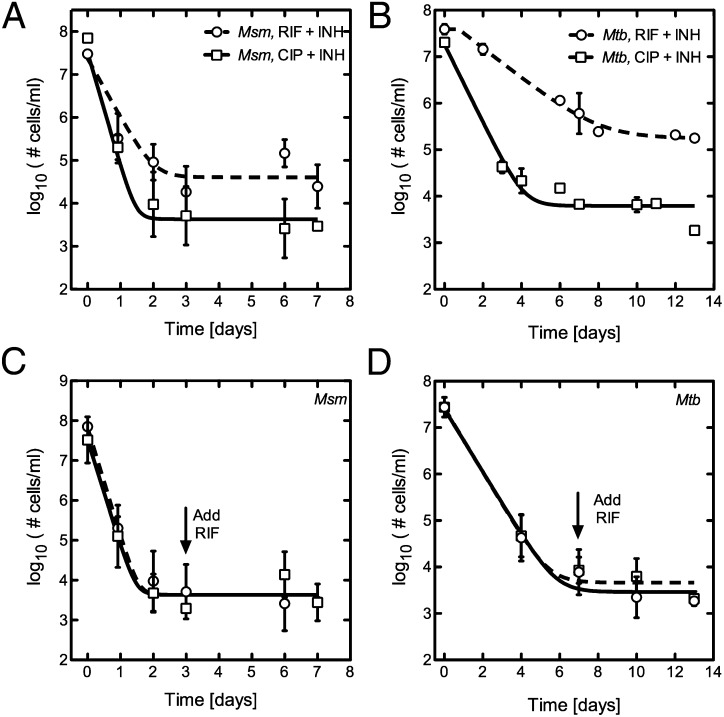
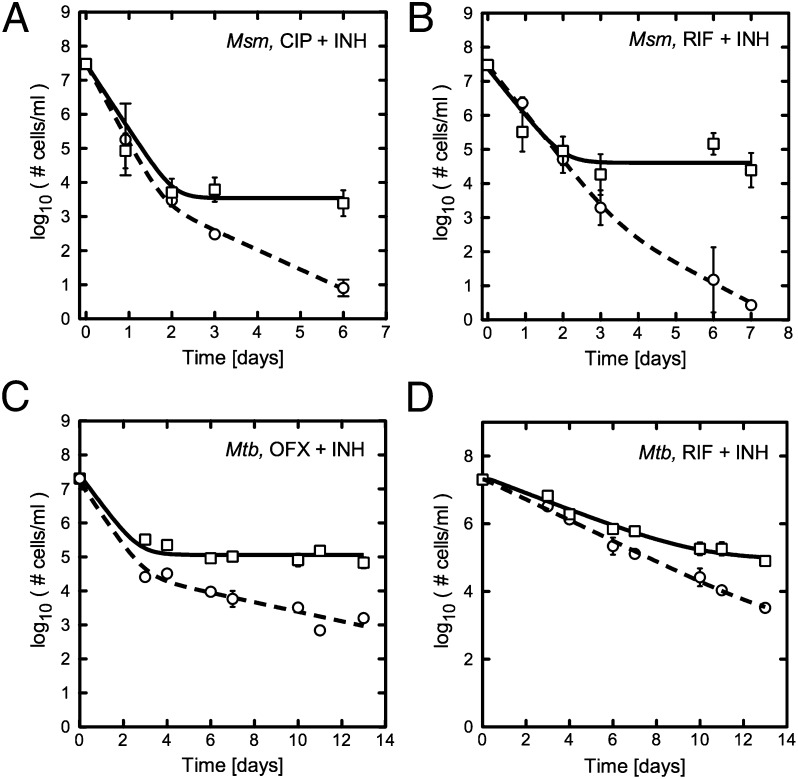
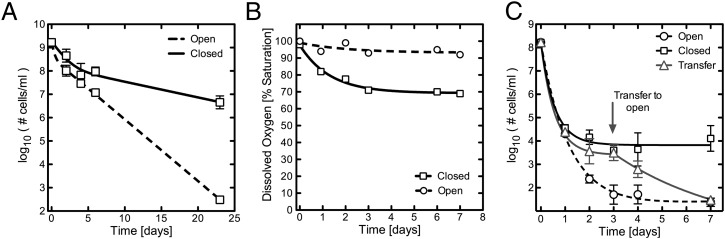
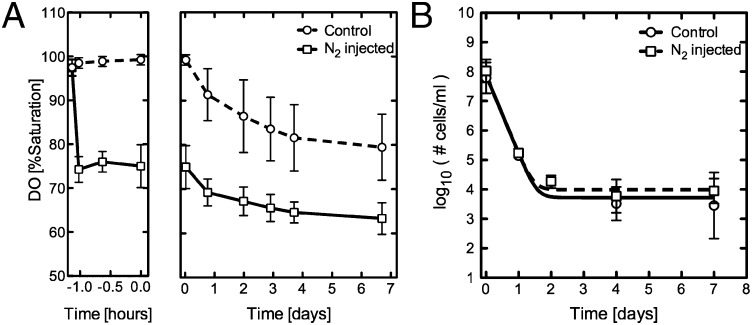
References
-
- Dye C, Scheele S, Dolin P, Pathania V, Raviglione MC. Consensus statement. Global burden of tuberculosis: Estimated incidence, prevalence, and mortality by country. WHO Global Surveillance and Monitoring Project. JAMA. 1999;282:677–686. - PubMed
-
- World Health Organization . TB Fact Sheet. Geneva: WHO; 2010.
-
- Sahbazian B, Weis SE. Treatment of active tuberculosis: Challenges and prospects. Clin Chest Med. 2005;26:273–282, vi. - PubMed
-
- Espinal MA. The global situation of MDR-TB. Tuberculosis. 2003;83:44–51. - PubMed
-
- Gomez JE, McKinney JD. M. tuberculosis persistence, latency, and drug tolerance. Tuberculosis. 2004;84:29–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
