

A designed ankyrin repeat protein selected to bind to tubulin caps the microtubule plus end
- PMID: 22778434
- PMCID: PMC3409770
- DOI: 10.1073/pnas.1204129109
A designed ankyrin repeat protein selected to bind to tubulin caps the microtubule plus end
Abstract
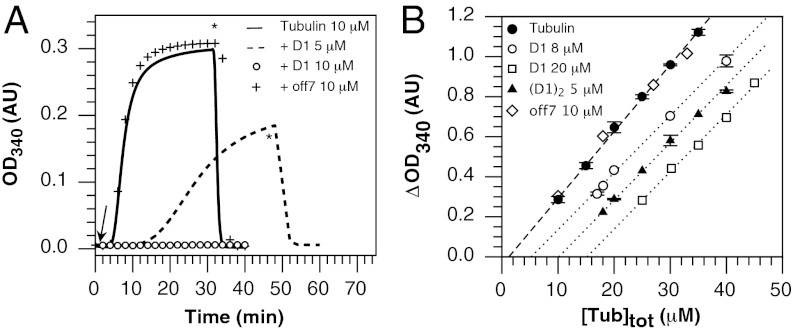
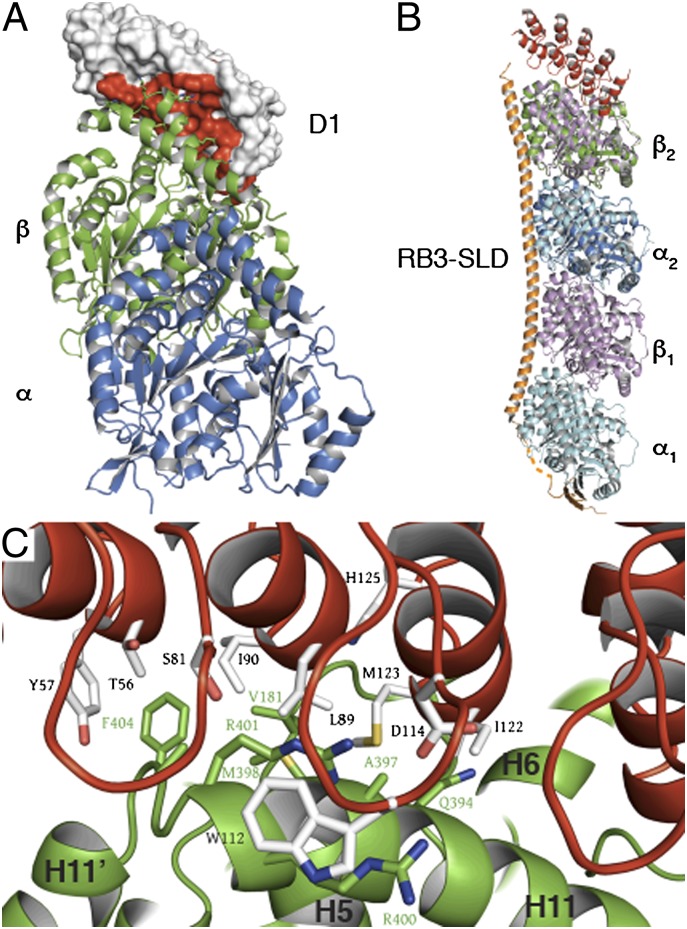
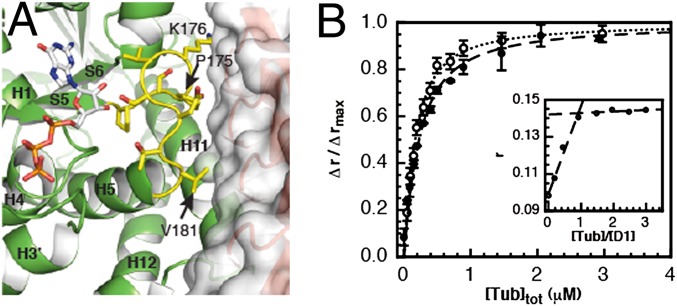
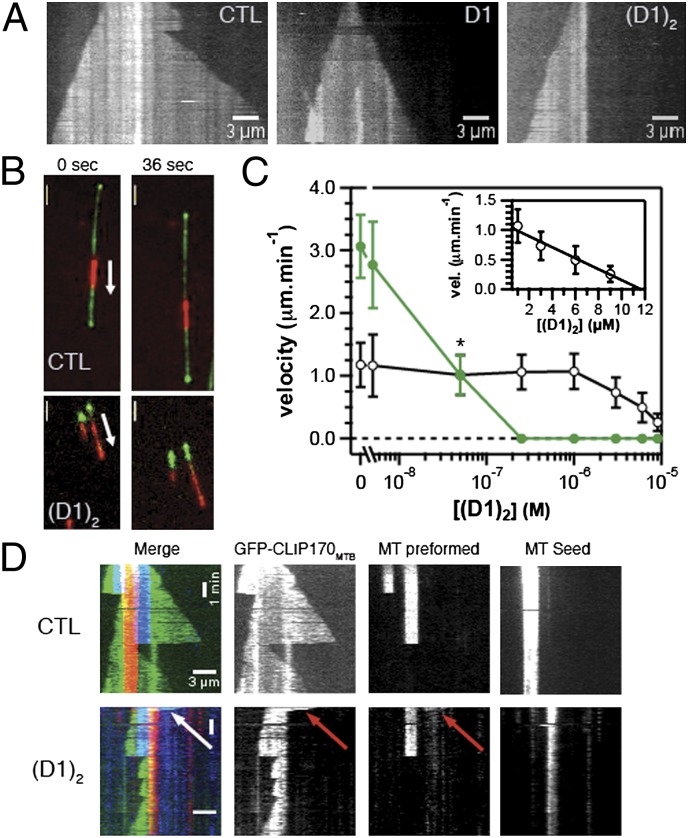
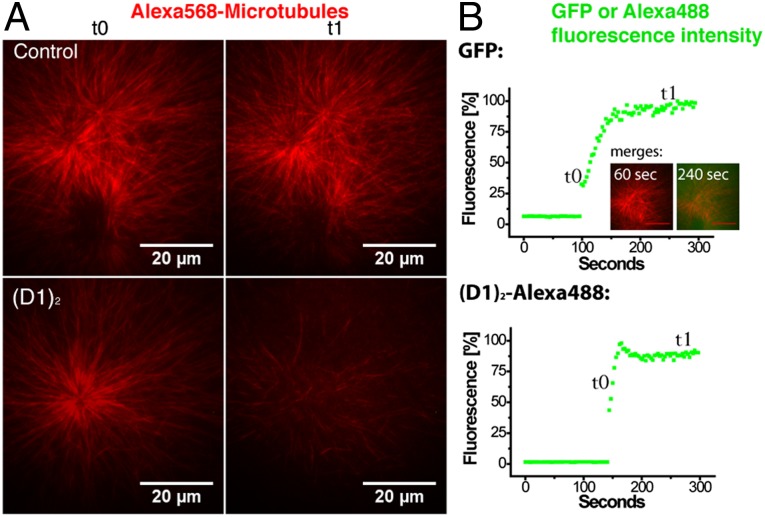
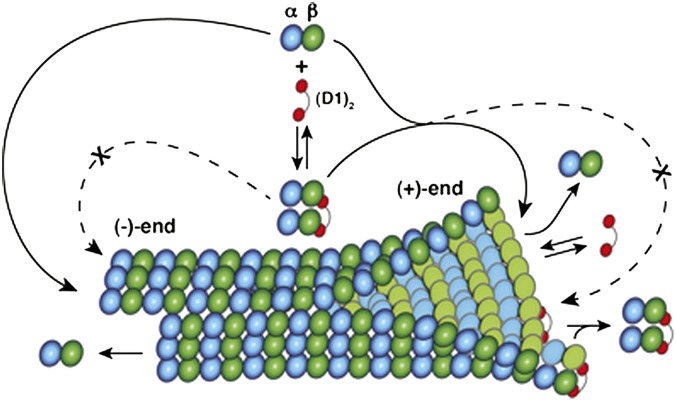
Microtubules are cytoskeleton filaments consisting of αβ-tubulin heterodimers. They switch between phases of growth and shrinkage. The underlying mechanism of this property, called dynamic instability, is not fully understood. Here, we identified a designed ankyrin repeat protein (DARPin) that interferes with microtubule assembly in a unique manner. The X-ray structure of its complex with GTP-tubulin shows that it binds to the β-tubulin surface exposed at microtubule (+) ends. The details of the structure provide insight into the role of GTP in microtubule polymerization and the conformational state of tubulin at the very microtubule end. They show in particular that GTP facilitates the tubulin structural switch that accompanies microtubule assembly but does not trigger it in unpolymerized tubulin. Total internal reflection fluorescence microscopy revealed that the DARPin specifically blocks growth at the microtubule (+) end by a selective end-capping mechanism, ultimately favoring microtubule disassembly from that end. DARPins promise to become designable tools for the dissection of microtubule dynamic properties selective for either of their two different ends.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
