

Reduced 3,4-methylenedioxymethamphetamine (MDMA, Ecstasy)-initiated oxidative DNA damage and neurodegeneration in prostaglandin H synthase-1 knockout mice
- PMID: 22778832
- PMCID: PMC3368639
- DOI: 10.1021/cn900022w
Reduced 3,4-methylenedioxymethamphetamine (MDMA, Ecstasy)-initiated oxidative DNA damage and neurodegeneration in prostaglandin H synthase-1 knockout mice
Abstract
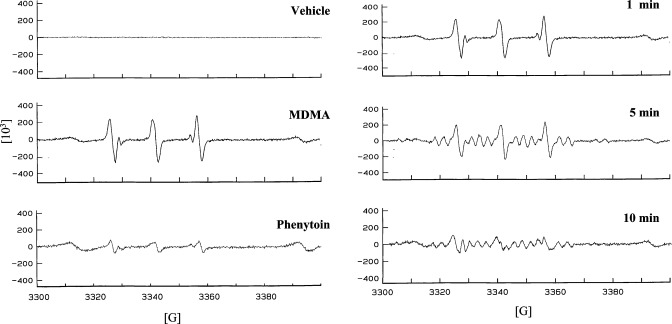
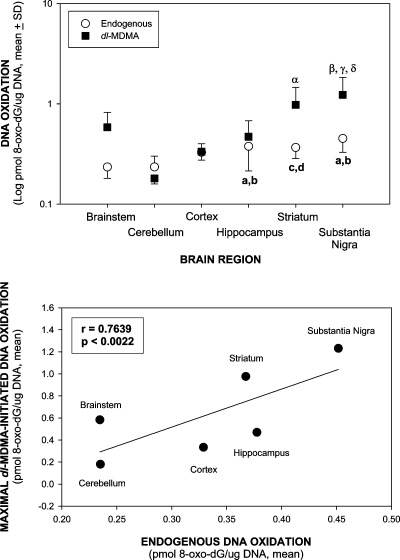
The neurodegenerative potential of 3,4-methylenedioxymethamphetamine (MDMA, ecstasy) and underlying mechanisms are under debate. Here, we show that MDMA is a substrate for CNS prostaglandin H synthase (PHS)-catalyzed bioactivation to a free radical intermediate that causes reactive oxygen species (ROS) formation and neurodegenerative oxidative DNA damage. In vitro PHS-1-catalyzed bioactivation of MDMA stereoselectively produced free radical intermediate formation and oxidative DNA damage that was blocked by the PHS inhibitor eicosatetraynoic acid. In vivo, MDMA stereoselectively caused gender-independent DNA oxidation and dopaminergic nerve terminal degeneration in several brain regions, dependent on regional PHS-1 levels. Conversely, MDMA-initiated striatal DNA oxidation, nerve terminal degeneration, and motor coordination deficits were reduced in PHS-1 +/- and -/- knockout mice in a gene dose-dependent fashion. These results confirm the neurodegenerative potential of MDMA and provide the first direct evidence for a novel molecular mechanism involving PHS-catalyzed formation of a neurotoxic MDMA free radical intermediate.
Keywords: 3,4-methylenedioxymethamphetamine; Ecstasy; neurodegeneration; oxidative DNA damage; prostaglandin H synthase; reactive oxygen species.
Figures






Similar articles
-
Prostaglandin H synthase-catalyzed bioactivation of amphetamines to free radical intermediates that cause CNS regional DNA oxidation and nerve terminal degeneration.FASEB J. 2006 Apr;20(6):638-50. doi: 10.1096/fj.05-5271com. FASEB J. 2006. PMID: 16581972
-
Reduced DNA oxidation in aged prostaglandin H synthase-1 knockout mice.Free Radic Biol Med. 2011 Feb 15;50(4):550-6. doi: 10.1016/j.freeradbiomed.2010.11.016. Epub 2010 Nov 18. Free Radic Biol Med. 2011. PMID: 21094252
-
A study of the mechanisms involved in the neurotoxic action of 3,4-methylenedioxymethamphetamine (MDMA, 'ecstasy') on dopamine neurones in mouse brain.Br J Pharmacol. 2001 Dec;134(8):1711-23. doi: 10.1038/sj.bjp.0704435. Br J Pharmacol. 2001. PMID: 11739248 Free PMC article.
-
The pharmacology and clinical pharmacology of 3,4-methylenedioxymethamphetamine (MDMA, "ecstasy").Pharmacol Rev. 2003 Sep;55(3):463-508. doi: 10.1124/pr.55.3.3. Epub 2003 Jul 17. Pharmacol Rev. 2003. PMID: 12869661 Review.
-
On the mechanisms underlying 3,4-methylenedioxymethamphetamine toxicity: the dilemma of the chicken and the egg.Neuropsychobiology. 2009;60(3-4):119-29. doi: 10.1159/000253548. Epub 2009 Nov 5. Neuropsychobiology. 2009. PMID: 19893329 Review.
Cited by
-
Investigations of the genotoxic properties of two synthetic cathinones (3-MMC, 4-MEC) which are used as psychoactive drugs.Toxicol Res (Camb). 2016 Jul 22;5(5):1410-1420. doi: 10.1039/c6tx00087h. eCollection 2016 Sep 1. Toxicol Res (Camb). 2016. PMID: 30090445 Free PMC article.
-
Prenatal Exposure to Methamphetamine: Up-Regulation of Brain Receptor Genes.Front Neurosci. 2019 Aug 1;13:771. doi: 10.3389/fnins.2019.00771. eCollection 2019. Front Neurosci. 2019. PMID: 31417344 Free PMC article.
-
Recent Advances on Electrochemical Sensors for Detection of Contaminants of Emerging Concern (CECs).Molecules. 2023 Dec 3;28(23):7916. doi: 10.3390/molecules28237916. Molecules. 2023. PMID: 38067644 Free PMC article. Review.
-
Brain glucose-6-phosphate dehydrogenase protects against endogenous oxidative DNA damage and neurodegeneration in aged mice.ACS Chem Neurosci. 2013 Jul 17;4(7):1123-32. doi: 10.1021/cn400079y. Epub 2013 May 14. ACS Chem Neurosci. 2013. PMID: 23672460 Free PMC article.
-
Breast cancer 1 (BRCA1)-deficient embryos develop normally but are more susceptible to ethanol-initiated DNA damage and embryopathies.Redox Biol. 2016 Apr;7:30-38. doi: 10.1016/j.redox.2015.11.005. Epub 2015 Nov 18. Redox Biol. 2016. PMID: 26629949 Free PMC article.
References
-
- Nichols D. E. (1986) Differences between the mechanism of action of MDMA, MBDB, and the classic hallucinogens. Identification of a new therapeutic class: entactogens. J. Psychoactive Drugs 18, 305–313. - PubMed
-
- Broening H. W.; Bowyer J. F.; Slikker W. (1995) Age-dependent sensitivity of rats to the long-term effects of the serotonergic neurotoxicant (±)-3,4-methylenedioxymethamphetamine (MDMA) correlates with the magnitude of the MDMA-induced thermal response. J. Pharmacol. Exp. Ther. 275, 325–333. - PubMed
-
- Bolla K. I.; McCann U. D.; Ricaurte G. A. (1998) Memory impairment in abstinent MDMA (“Ecstasy”) users. Neurology 51, 1532–1537. - PubMed
-
- McGuire P. (2000) Long term psychiatric and cognitive effects of MDMA use. Toxicol. Lett. 112−113, 153–156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
