

Clustered metallothionein genes are co-regulated in rice and ectopic expression of OsMT1e-P confers multiple abiotic stress tolerance in tobacco via ROS scavenging
- PMID: 22780875
- PMCID: PMC3491035
- DOI: 10.1186/1471-2229-12-107
Clustered metallothionein genes are co-regulated in rice and ectopic expression of OsMT1e-P confers multiple abiotic stress tolerance in tobacco via ROS scavenging
Abstract
Background: Metallothioneins (MT) are low molecular weight, cysteine rich metal binding proteins, found across genera and species, but their function(s) in abiotic stress tolerance are not well documented.
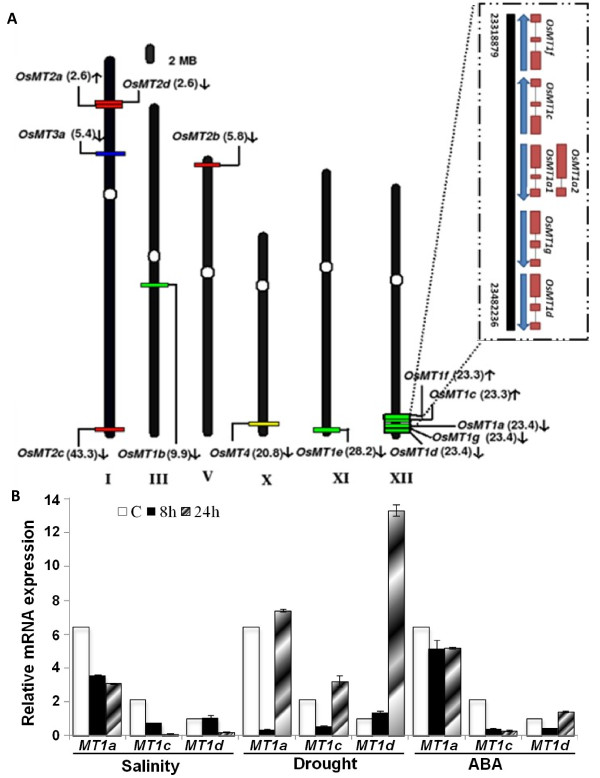
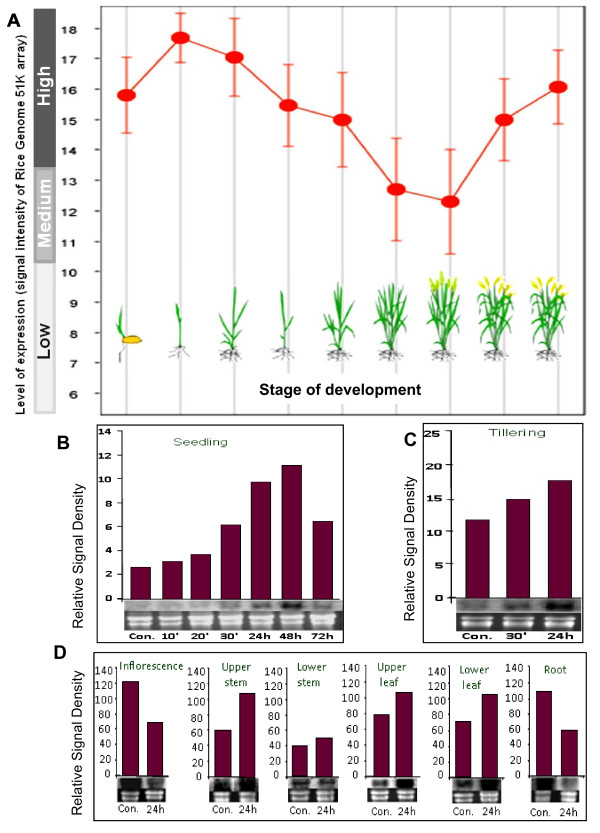
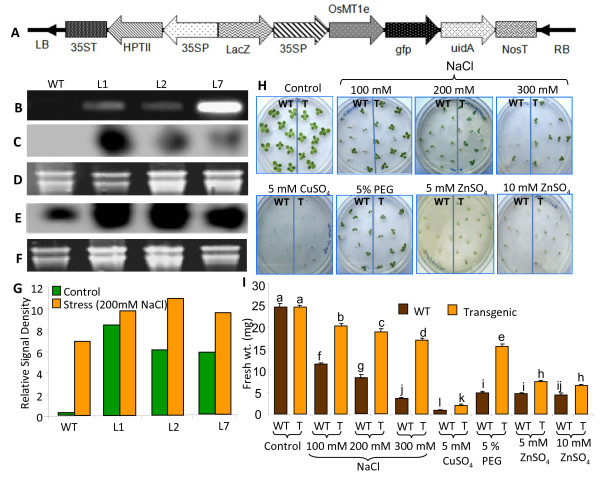
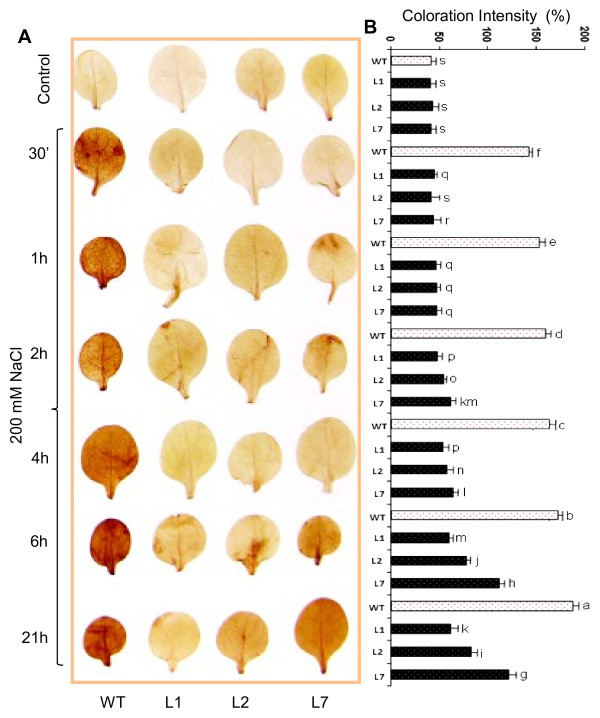
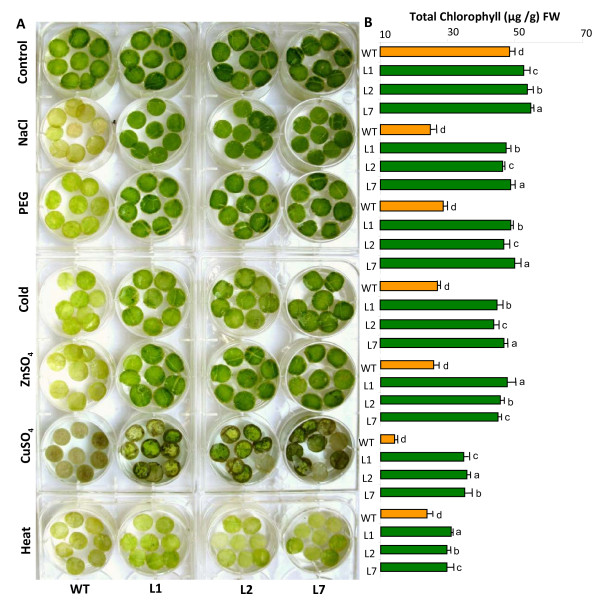
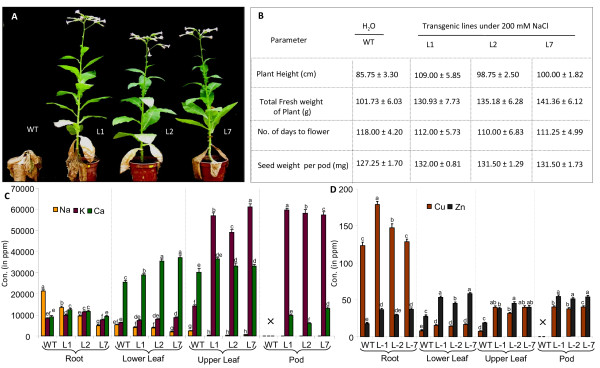
Results: We have characterized a rice MT gene, OsMT1e-P, isolated from a subtractive library generated from a stressed salinity tolerant rice genotype, Pokkali. Bioinformatics analysis of the rice genome sequence revealed that this gene belongs to a multigenic family, which consists of 13 genes with 15 protein products. OsMT1e-P is located on chromosome XI, away from the majority of other type I genes that are clustered on chromosome XII. Various members of this MT gene cluster showed a tight co-regulation pattern under several abiotic stresses. Sequence analysis revealed the presence of conserved cysteine residues in OsMT1e-P protein. Salinity stress was found to regulate the transcript abundance of OsMT1e-P in a developmental and organ specific manner. Using transgenic approach, we found a positive correlation between ectopic expression of OsMT1e-P and stress tolerance. Our experiments further suggest ROS scavenging to be the possible mechanism for multiple stress tolerance conferred by OsMT1e-P.
Conclusion: We present an overview of MTs, describing their gene structure, genome localization and expression patterns under salinity and development in rice. We have found that ectopic expression of OsMT1e-P enhances tolerance towards multiple abiotic stresses in transgenic tobacco and the resultant plants could survive and set viable seeds under saline conditions. Taken together, the experiments presented here have indicated that ectopic expression of OsMT1e-P protects against oxidative stress primarily through efficient scavenging of reactive oxygen species.
Figures
References
-
- Cobbett CS, Goldsbrough P. Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Ann Rev Plant Physiol Mol Biol. 2002;53:159–182. - PubMed
-
- Cherian GM, Chan HM. In: Metallothionein III: Biological Roles and Medical Implications. Suzuki KT, Imura N, Kimura M, editor. Birkhauser Verlag, Boston; 1993. Biological functions of metallothioneins-a review; pp. 87–109.
-
- Binz PA, Kagi JHR. In: Metallothionein IV. Klaassen C, editor. Basel, Switzerland: Birkhauser Verlag; 1999. Metallothionein: molecular evolution and classification; pp. 7–13.
-
- Freisinger E. Metallothioneins in plants. Met Ions Life Sci. 2009;5:107–153. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
