

Mycobacterium tuberculosis WhiB4 regulates oxidative stress response to modulate survival and dissemination in vivo
- PMID: 22780904
- PMCID: PMC3438311
- DOI: 10.1111/j.1365-2958.2012.08165.x
Mycobacterium tuberculosis WhiB4 regulates oxidative stress response to modulate survival and dissemination in vivo
Abstract
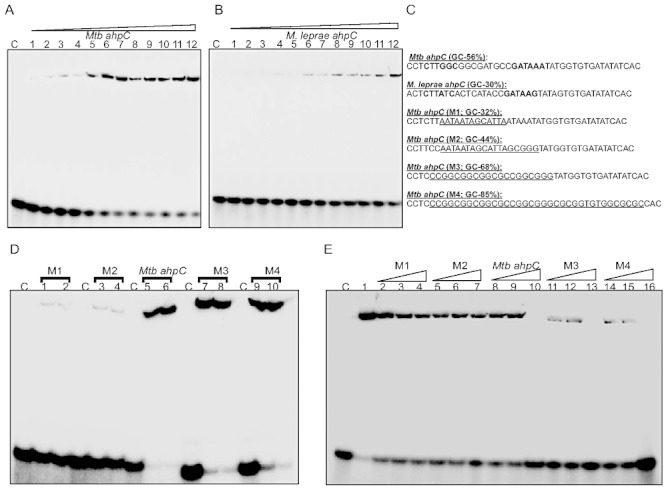
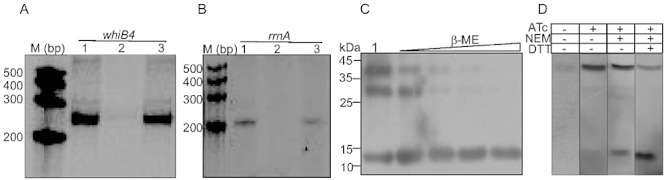
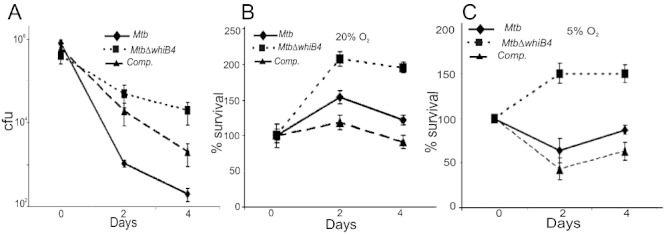
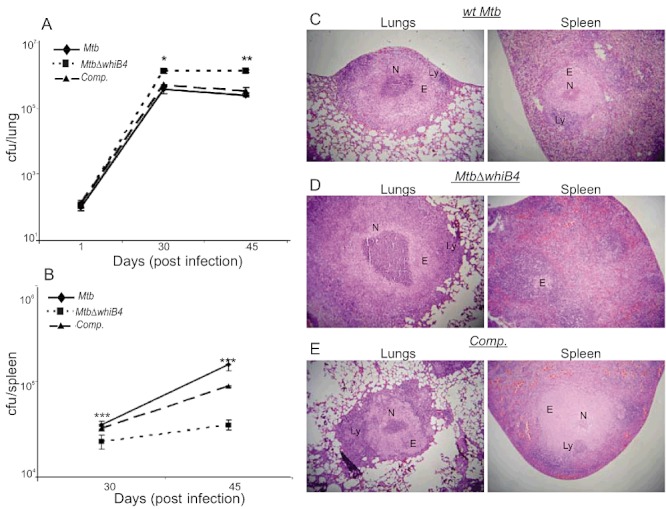
Host-generated oxidative stress is considered one of the main mechanisms constraining Mycobacterium tuberculosis (Mtb) growth. The redox-sensing mechanisms in Mtb are not completely understood. Here we show that WhiB4 responds to oxygen (O₂) and nitric oxide (NO) via its 4Fe-4S cluster and controls the oxidative stress response in Mtb. The WhiB4 mutant (MtbΔwhiB4) displayed an altered redox balance and a reduced membrane potential. Microarray analysis demonstrated that MtbΔwhiB4 overexpresses the antioxidant systems including alkyl hydroperoxidase (ahpC-ahpD) and rubredoxins (rubA-rubB). DNA binding assays showed that WhiB4 [4Fe-4S] cluster is dispensable for DNA binding. However, oxidation of the apo-WhiB4 Cys thiols induced disulphide-linked oligomerization, DNA binding and transcriptional repression, whereas reduction reversed the effect. Furthermore, WhiB4 binds DNA with a preference for GC-rich sequences. Expression analysis showed that oxidative stress repressed whiB4 and induced antioxidants in Mtb, while their hyper-induction was observed in MtbΔwhiB4. MtbΔwhiB4 showed increased resistance to oxidative stress in vitro and enhanced survival inside the macrophages. Lastly, MtbΔwhiB4 displayed hypervirulence in the lungs of guinea pigs, but showed a defect in dissemination to their spleen. These findings suggest that WhiB4 systematically calibrates the activation of oxidative stress response in Mtb to maintain redox balance, and to modulate virulence.
© 2012 Blackwell Publishing Ltd.
Figures
Similar articles
-
Redox-dependent condensation of the mycobacterial nucleoid by WhiB4.Redox Biol. 2018 Oct;19:116-133. doi: 10.1016/j.redox.2018.08.006. Epub 2018 Aug 13. Redox Biol. 2018. PMID: 30149290 Free PMC article.
-
Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response.PLoS Pathog. 2009 Aug;5(8):e1000545. doi: 10.1371/journal.ppat.1000545. Epub 2009 Aug 14. PLoS Pathog. 2009. PMID: 19680450 Free PMC article.
-
Mycobacterium tuberculosis EsxO (Rv2346c) promotes bacillary survival by inducing oxidative stress mediated genomic instability in macrophages.Tuberculosis (Edinb). 2016 Jan;96:44-57. doi: 10.1016/j.tube.2015.11.006. Epub 2015 Nov 24. Tuberculosis (Edinb). 2016. PMID: 26786654
-
Redox biology of tuberculosis pathogenesis.Adv Microb Physiol. 2012;60:263-324. doi: 10.1016/B978-0-12-398264-3.00004-8. Adv Microb Physiol. 2012. PMID: 22633061 Review.
-
Deciphering the role of VapBC toxin-antitoxin systems in Mycobacterium tuberculosis stress adaptation.Future Microbiol. 2024;19(18):1587-1599. doi: 10.1080/17460913.2024.2412447. Epub 2024 Oct 21. Future Microbiol. 2024. PMID: 39431307 Review.
Cited by
-
Vanoxerine kills mycobacteria through membrane depolarization and efflux inhibition.Front Microbiol. 2023 Jan 26;14:1112491. doi: 10.3389/fmicb.2023.1112491. eCollection 2023. Front Microbiol. 2023. PMID: 36778873 Free PMC article.
-
An intricate regulation of WblA controlling production of silent tylosin analogues and abolishment of expressible nikkomycin.Sci China Life Sci. 2023 Mar;66(3):612-625. doi: 10.1007/s11427-022-2199-1. Epub 2023 Jan 4. Sci China Life Sci. 2023. PMID: 36607495
-
Mycobacterium tuberculosis requires SufT for Fe-S cluster maturation, metabolism, and survival in vivo.PLoS Pathog. 2022 Apr 15;18(4):e1010475. doi: 10.1371/journal.ppat.1010475. eCollection 2022 Apr. PLoS Pathog. 2022. PMID: 35427399 Free PMC article.
-
Cysteine desulfurase (IscS)-mediated fine-tuning of bioenergetics and SUF expression prevents Mycobacterium tuberculosis hypervirulence.Sci Adv. 2023 Dec 15;9(50):eadh2858. doi: 10.1126/sciadv.adh2858. Epub 2023 Dec 13. Sci Adv. 2023. PMID: 38091389 Free PMC article.
-
Reengineering redox sensitive GFP to measure mycothiol redox potential of Mycobacterium tuberculosis during infection.PLoS Pathog. 2014 Jan 30;10(1):e1003902. doi: 10.1371/journal.ppat.1003902. eCollection 2014 Jan. PLoS Pathog. 2014. PMID: 24497832 Free PMC article.
References
-
- Alam MS, Garg SK, Agrawal P. Molecular function of WhiB4/Rv3681c of Mycobacterium tuberculosis H37Rv: a [4Fe-4S] cluster co-ordinating protein disulphide reductase. Mol Microbiol. 2007;63:1414–1431. - PubMed
-
- Alam MS, Garg SK, Agrawal P. Studies on structural and functional divergence among seven WhiB proteins of Mycobacterium tuberculosis H37Rv. FEBS J. 2009;276:76–93. - PubMed
-
- Bardarov S, Bardarov S, Jr, Pavelka MS, Jr, Sambandamurthy V, Larsen M, Tufariello J, et al. Specialized transduction: an efficient method for generating marked and unmarked targeted gene disruptions in Mycobacterium tuberculosis, M. bovis BCG and M. smegmatis. Microbiology. 2002;148:3007–3017. - PubMed
-
- Betts JC, Lukey PT, Robb LC, McAdam RA, Duncan K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol Microbiol. 2002;43:717–731. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
