

Lipoprotein lipase is frequently overexpressed or translocated in cervical squamous cell carcinoma and promotes invasiveness through the non-catalytic C terminus
- PMID: 22782350
- PMCID: PMC3419954
- DOI: 10.1038/bjc.2012.301
Lipoprotein lipase is frequently overexpressed or translocated in cervical squamous cell carcinoma and promotes invasiveness through the non-catalytic C terminus
Abstract
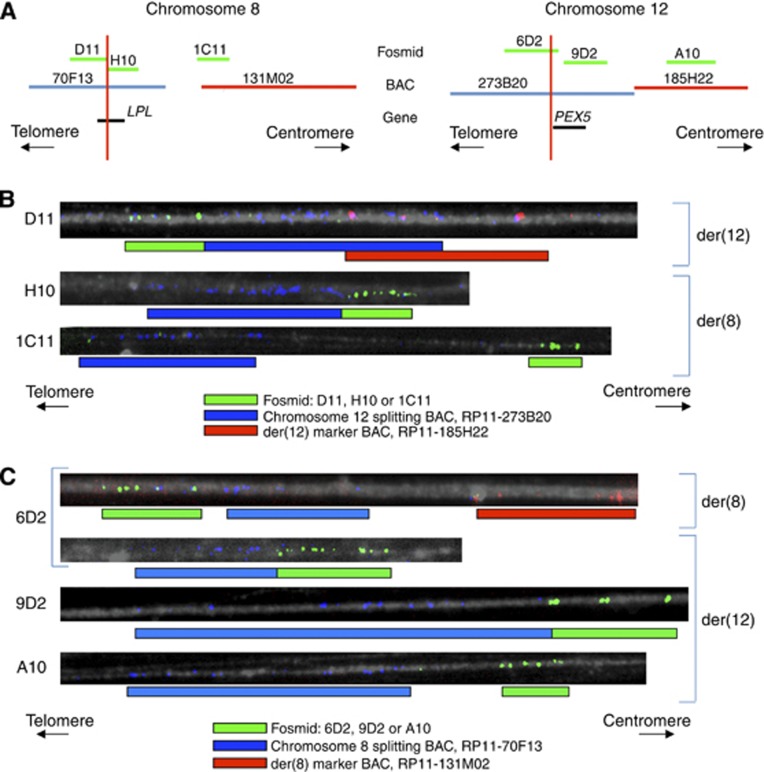
Background: We studied the biological significance of genes involved in a novel t(8;12)(p21.3;p13.31) reciprocal translocation identified in cervical squamous cell carcinoma (SCC) cells.
Methods: The rearranged genes were identified by breakpoint mapping, long-range PCR and sequencing. We investigated gene expression in vivo using reverse-transcription PCR and tissue microarrays, and studied the phenotypic consequences of forced gene overexpression.
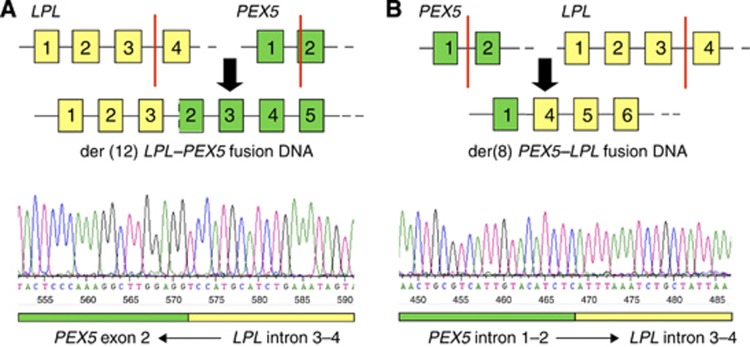
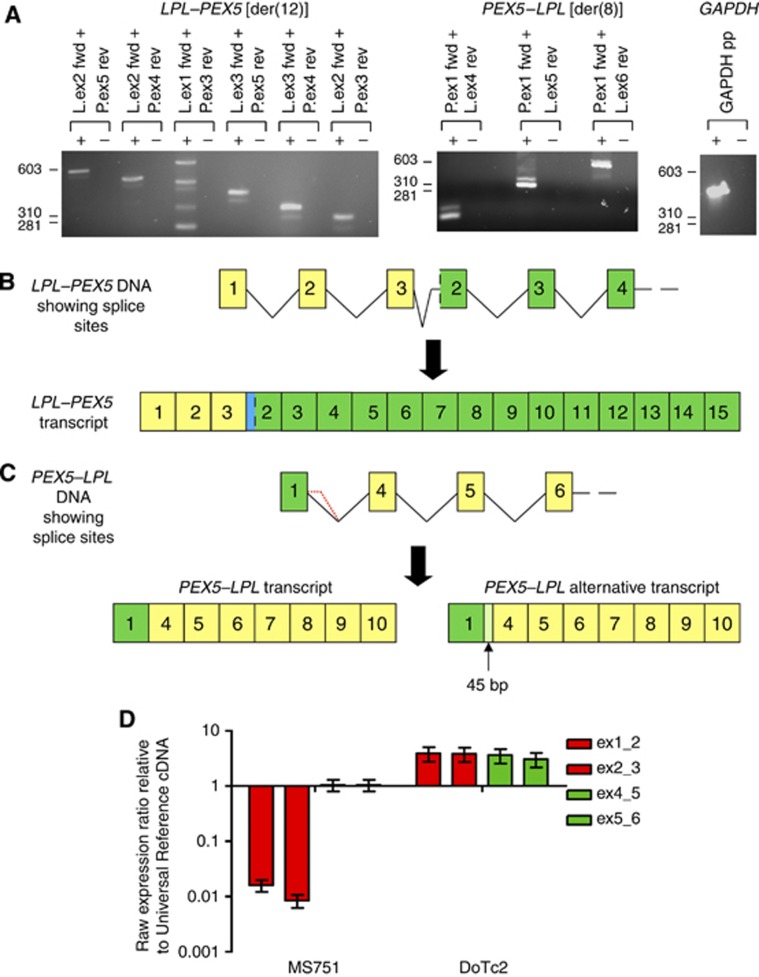
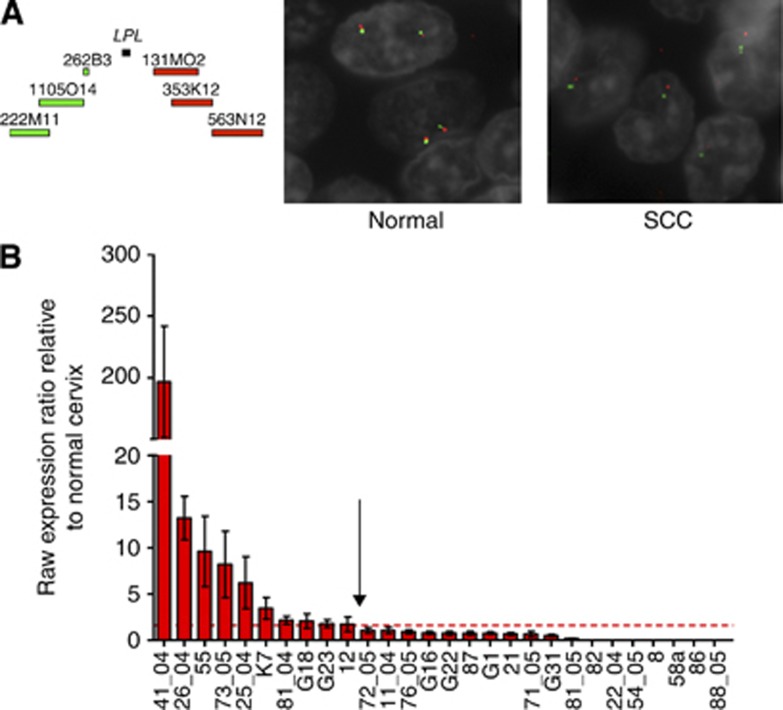
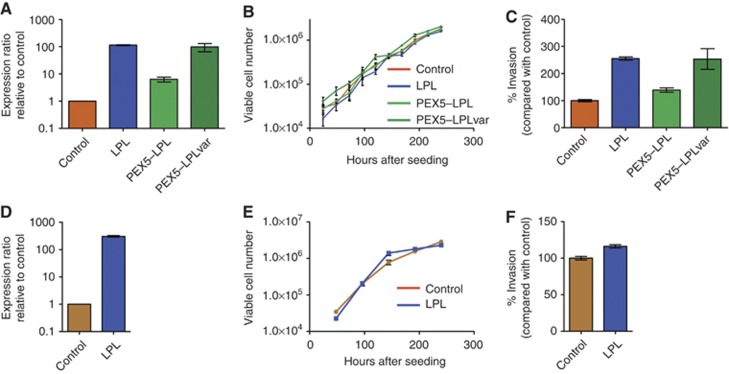
Results: The rearrangement involved lipoprotein lipase (LPL) and peroxisome biogenesis factor-5 (PEX5). Whereas LPL-PEX5 was expressed at low levels and contained a premature stop codon, PEX5-LPL was highly expressed and encoded a full-length chimeric protein (including the majority of the LPL coding region). Consistent with these findings, PEX5 was constitutively expressed in normal cervical squamous cells, whereas LPL expression was negligible. The LPL gene was rearranged in 1 out of 151 cervical SCCs, whereas wild-type LPL overexpression was common, being detected in 10 out of 28 tissue samples and 4 out of 10 cell lines. Forced overexpression of wild-type LPL and PEX5-LPL fusion transcripts resulted in increased invasiveness in cervical SCC cells, attributable to the C-terminal non-catalytic domain of LPL, which was retained in the fusion transcripts.
Conclusion: This is the first demonstration of an expressed fusion gene in cervical SCC. Overexpressed wild-type or translocated LPL is a candidate for targeted therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Baldwin P, Laskey R, Coleman N (2003) Translational approaches to improving cervical screening. Nat Rev Cancer 3(3): 217–226 - PubMed
-
- Ben-Zeev O, Stahnke G, Liu G, Davis RC, Doolittle MH (1994) Lipoprotein lipase and hepatic lipase: the role of asparagine-linked glycosylation in the expression of a functional enzyme. J Lipid Res 35(9): 1511–1523 - PubMed
-
- Blanchette-Mackie EJ, Masuno H, Dwyer NK, Olivecrona T, Scow RO (1989) Lipoprotein lipase in myocytes and capillary endothelium of heart: immunocytochemical study. Am J Physiol 256(6 Part 1): E818–E828 - PubMed
-
- Bosch FX, Manos MM, Munoz N, Sherman M, Jansen AM, Peto J, Schiffman MH, Moreno V, Kurman R, Shah KV (1995) Prevalence of human papillomavirus in cervical cancer: a worldwide perspective. International biological study on cervical cancer (IBSCC) Study Group. J Natl Cancer Inst 87(11): 796–802 - PubMed
