

Deciphering the role of CD1e protein in mycobacterial phosphatidyl-myo-inositol mannosides (PIM) processing for presentation by CD1b to T lymphocytes
- PMID: 22782895
- PMCID: PMC3438982
- DOI: 10.1074/jbc.M112.386300
Deciphering the role of CD1e protein in mycobacterial phosphatidyl-myo-inositol mannosides (PIM) processing for presentation by CD1b to T lymphocytes
Abstract
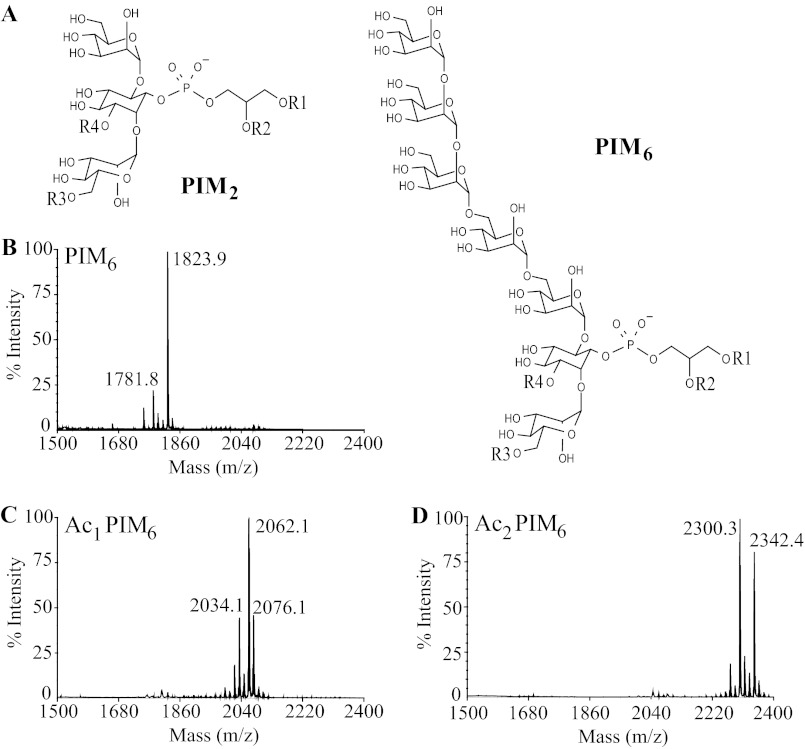
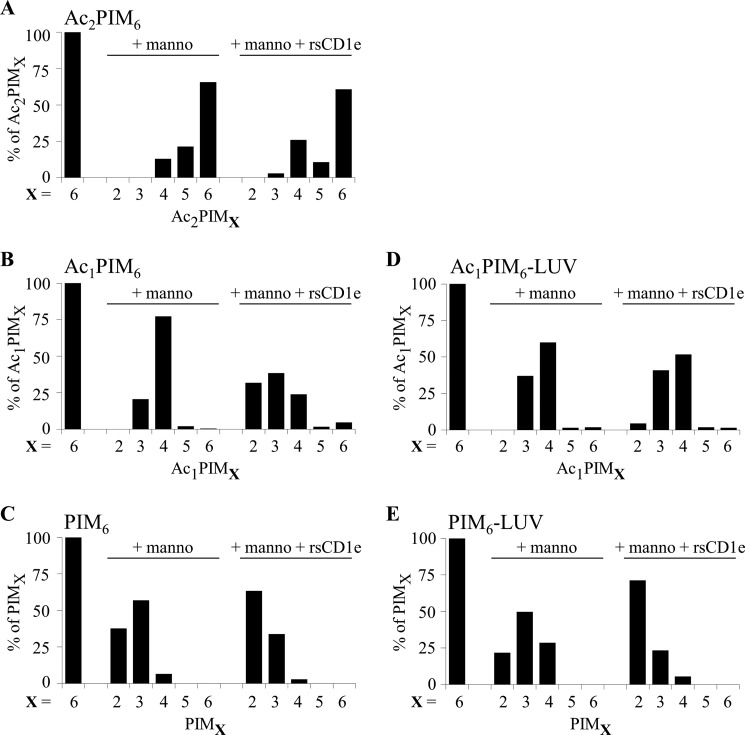
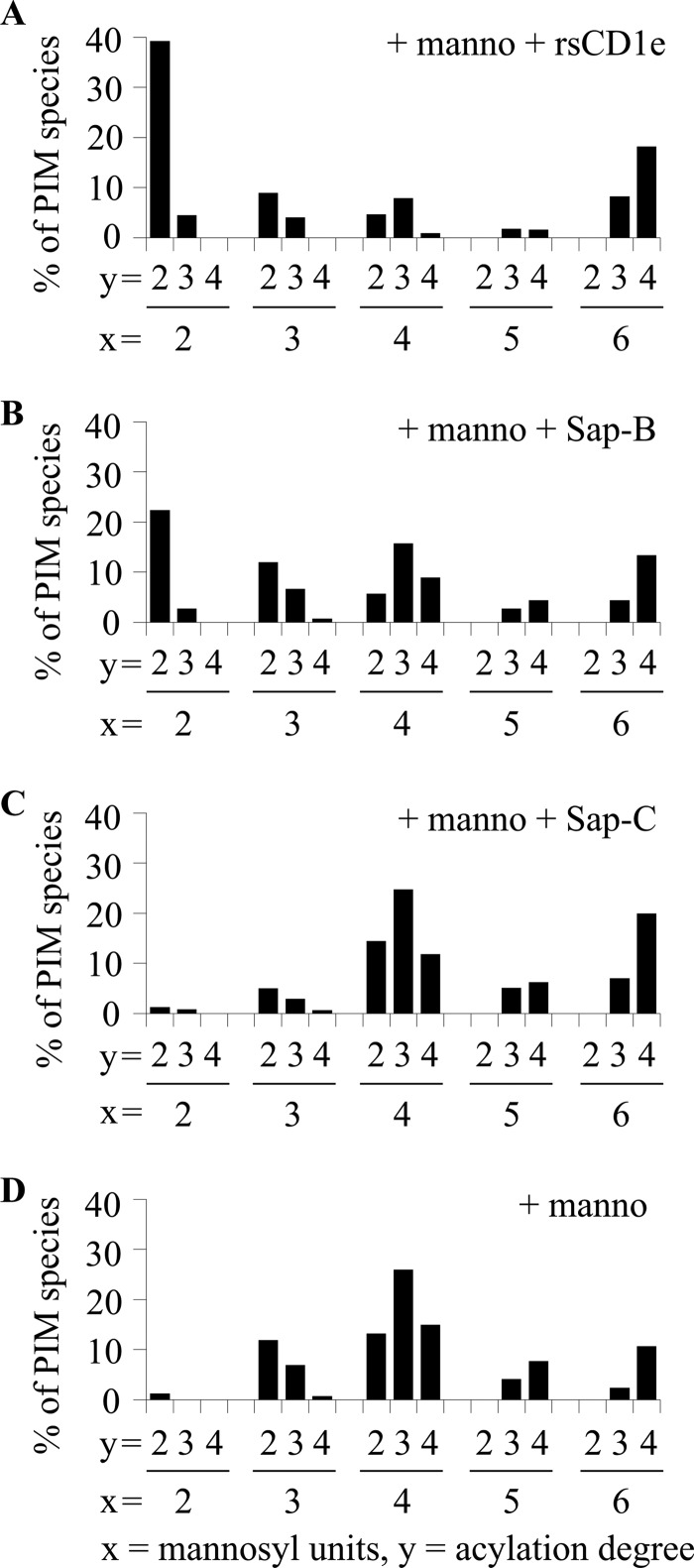
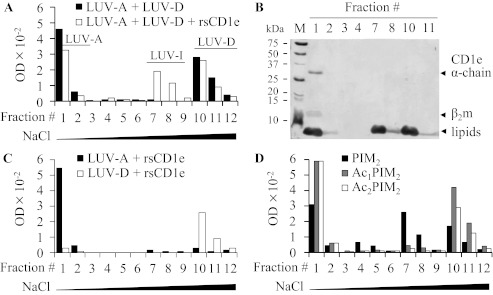
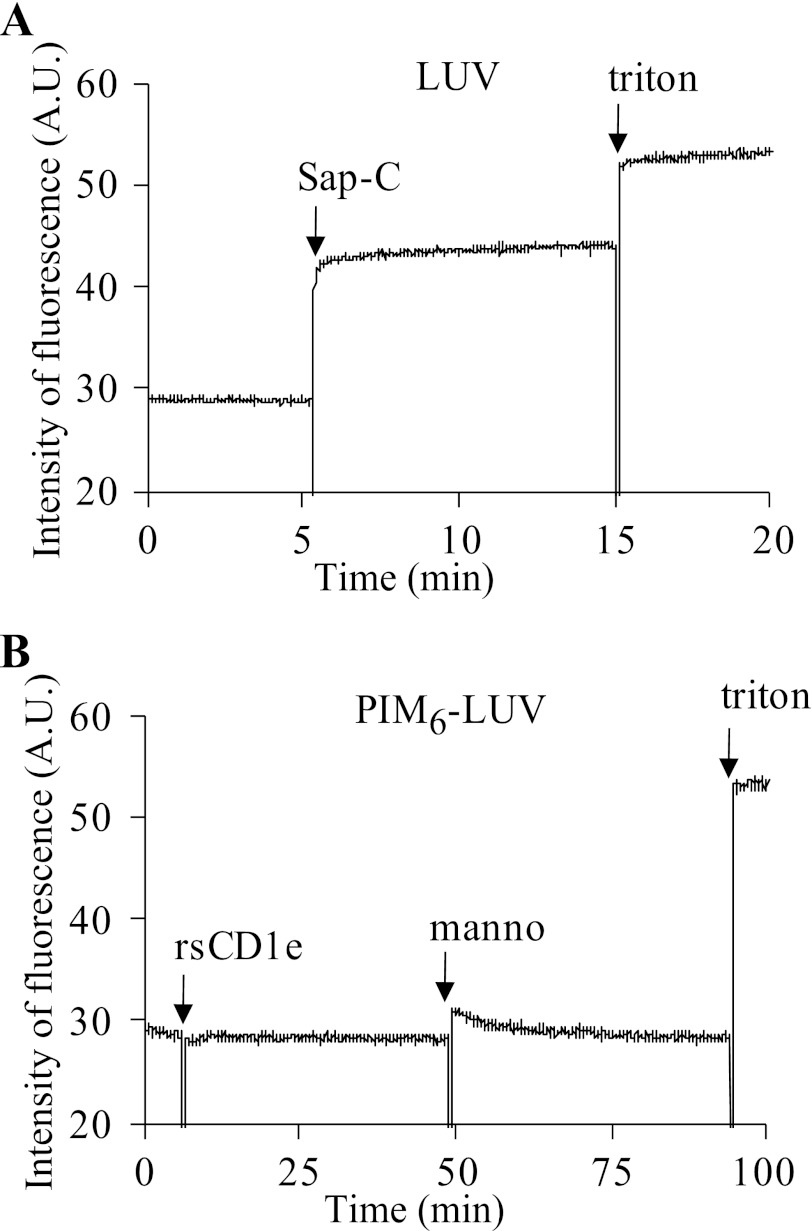
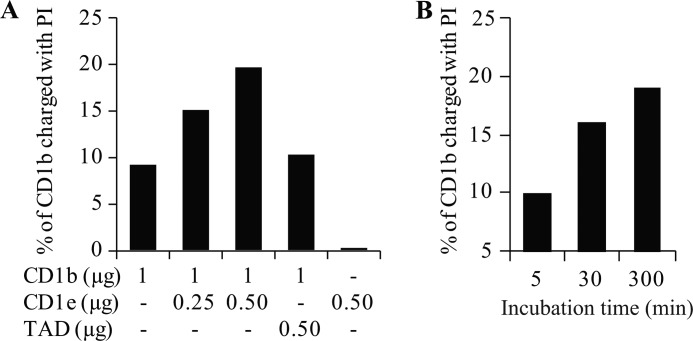
Lipids are important antigens that induce T cell-mediated specific immune responses. They are presented to T lymphocytes by a specific class of MHC-I like proteins, termed CD1. The majority of the described CD1-presented mycobacterial antigens are presented by the CD1b isoform. We previously demonstrated that the stimulation of CD1b-restricted T cells by the hexamannosylated phosphatidyl-myo-inositol (PIM(6)), a family of mycobacterial antigens, requires a prior partial digestion of the antigen oligomannoside moiety by α-mannosidase and that CD1e is an accessory protein absolutely required for the generation of the lipid immunogenic form. Here, we show that CD1e behaves as a lipid transfer protein influencing lipid immunoediting and membrane transfer of PIM lipids. CD1e selectively assists the α-mannosidase-dependent digestion of PIM(6) species according to their degree of acylation. Moreover, CD1e transfers only diacylated PIM from donor to acceptor liposomes and also from membranes to CD1b. This study provides new insight into the molecular mechanisms by which CD1e contributes to lipid immunoediting and CD1-restricted presentation to T cells.
Figures
References
-
- Barral D. C., Brenner M. B. (2007) CD1 antigen presentation: How it works. Nat. Rev. Immunol. 7, 929–941 - PubMed
-
- De Libero G., Collmann A., Mori L. (2009) The cellular and biochemical rules of lipid antigen presentation. Eur. J. Immunol. 39, 2648–2656 - PubMed
-
- Dougan S. K., Kaser A., Blumberg R. S. (2007) CD1 expression on antigen-presenting cells. Curr. Top Microbiol. Immunol. 314, 113–141 - PubMed
-
- Sugita M., Grant E. P., van Donselaar E., Hsu V. W., Rogers R. A., Peters P. J., Brenner M. B. (1999) Separate pathways for antigen presentation by CD1 molecules. Immunity 11, 743–752 - PubMed
-
- Angenieux C., Salamero J., Fricker D., Cazenave J. P., Goud B., Hanau D., de La Salle H. (2000) Characterization of CD1e, a third type of CD1 molecule expressed in dendritic cells. J. Biol. Chem. 275, 37757–37764 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
