

A retroelement modifies pre-mRNA splicing: the murine Glrb(spa) allele is a splicing signal polymorphism amplified by long interspersed nuclear element insertion
- PMID: 22782896
- PMCID: PMC3438950
- DOI: 10.1074/jbc.M112.375691
A retroelement modifies pre-mRNA splicing: the murine Glrb(spa) allele is a splicing signal polymorphism amplified by long interspersed nuclear element insertion
Abstract
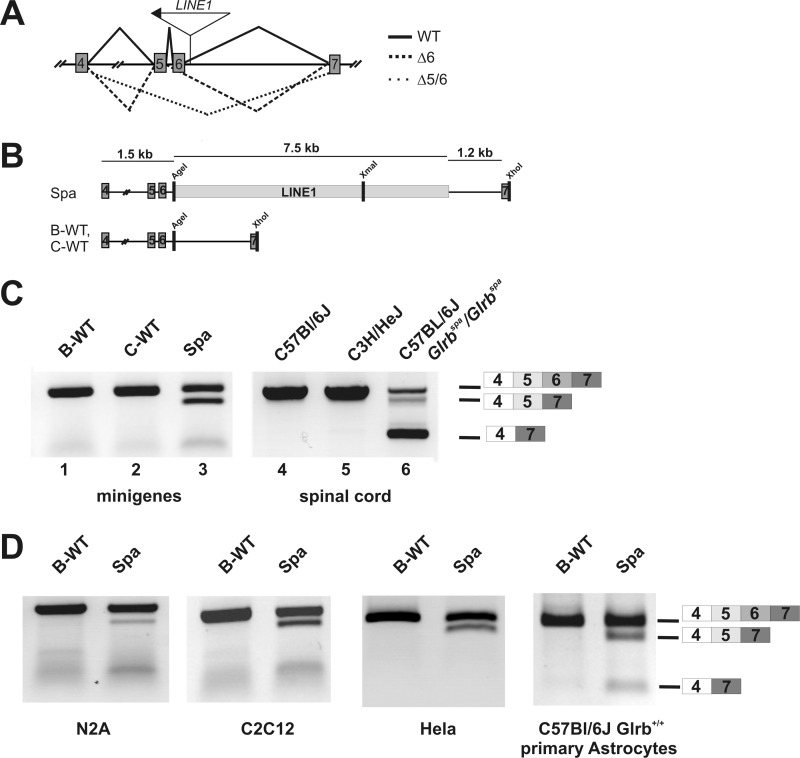
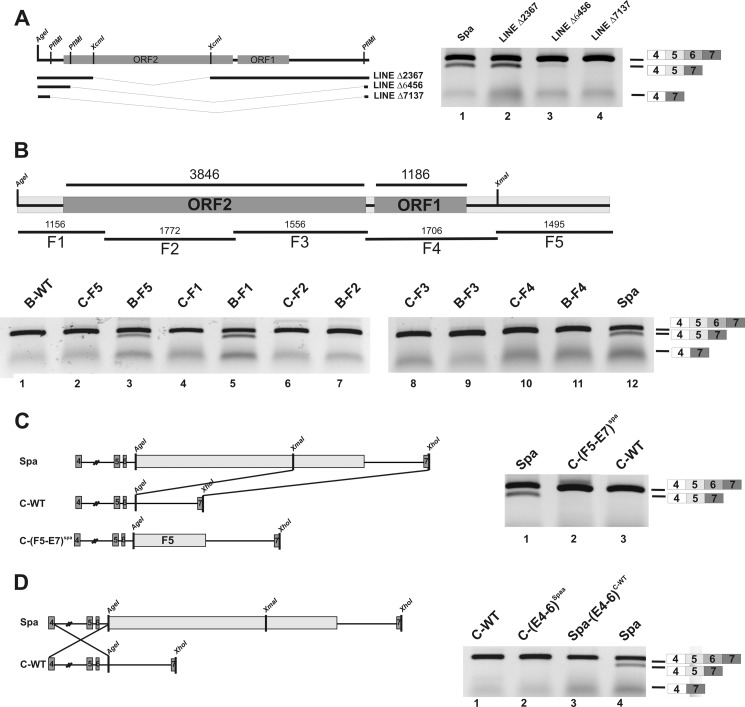
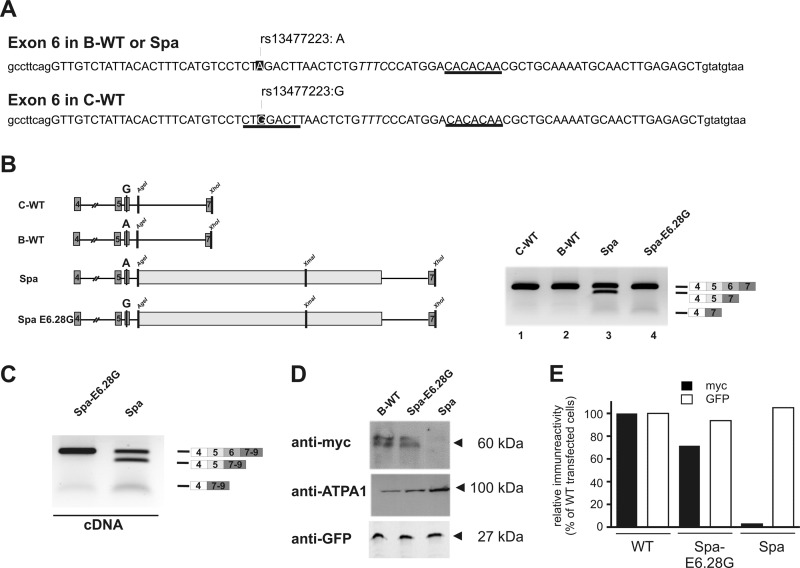
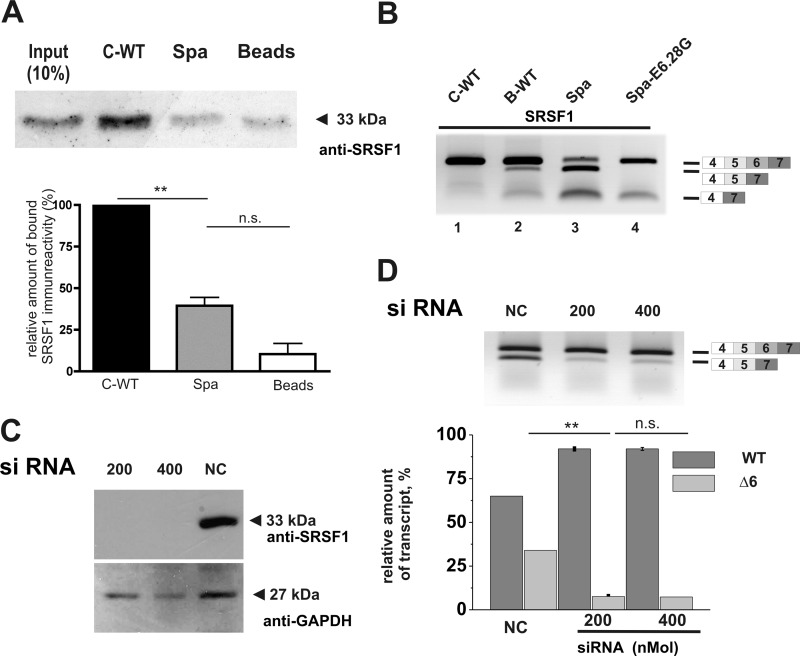
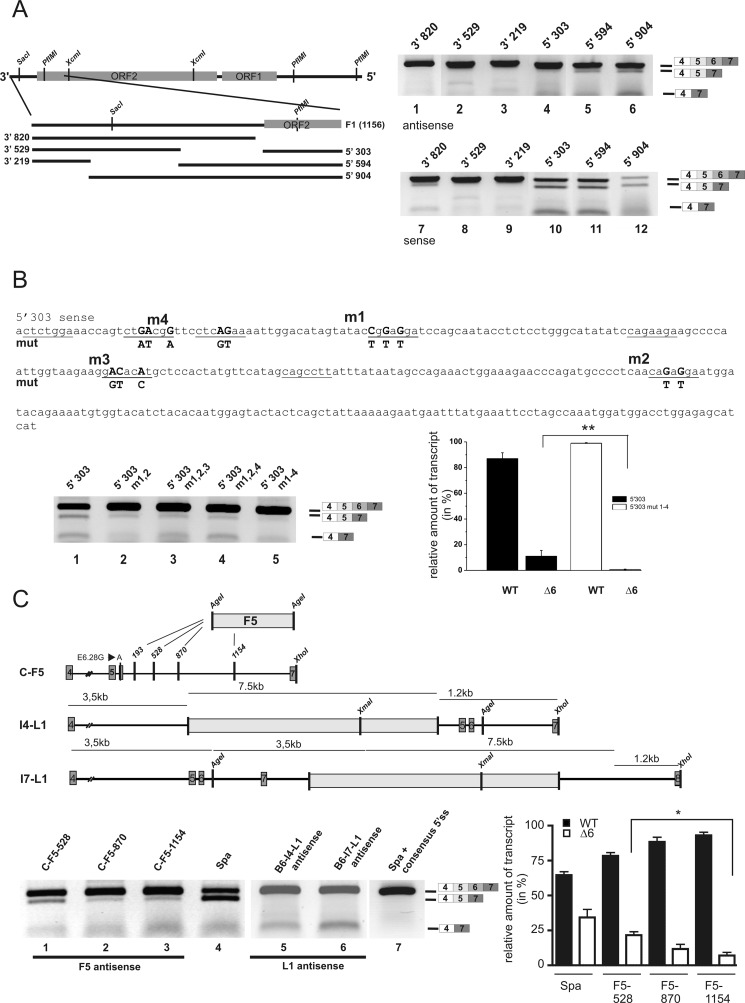
The glycine receptor-deficient mutant mouse spastic carries a full-length long interspersed nuclear element (LINE1) retrotransposon in intron 6 of the glycine receptor β subunit gene, Glrb(spa). The mutation arose in the C57BL/6J strain and is associated with skipping of exon 6 or a combination of the exons 5 and 6, thus resulting in a translational frameshift within the coding regions of the GlyR β subunit. The effect of the Glrb(spa) LINE1 insertion on pre-mRNA splicing was studied using a minigene approach. Sequence comparison as well as motif prediction and mutational analysis revealed that in addition to the LINE1 insertion the inactivation of an exonic splicing enhancer (ESE) within exon 6 is required for skipping of exon 6. Reconstitution of the ESE by substitution of a single residue was sufficient to prevent exon skipping. In addition to the ESE, two regions within the 5' and 3' UTR of the LINE1 were shown to be critical determinants for exon skipping, indicating that LINE1 acts as efficient modifier of subtle endogenous splicing phenotypes. Thus, the spastic allele of the murine glycine receptor β subunit gene is a two-hit mutation, where the hypomorphic alteration in an ESE is amplified by the insertion of a LINE1 element in the adjacent intron. Conversely, the LINE1 effect on splicing may be modulated by individual polymorphisms, depending on the insertional environment within the host genome.
Figures
References
-
- Betz H., Laube B. (2006) Glycine receptors: Recent insights into their structural organization and functional diversity. J. Neurochem. 97, 1600–1610 - PubMed
-
- Grudzinska J., Schemm R., Haeger S., Nicke A., Schmalzing G., Betz H., Laube B. (2005) The β subunit determines the ligand binding properties of synaptic glycine receptors. Neuron 45, 727–739 - PubMed
-
- Oertel J., Villmann C., Kettenmann H., Kirchhoff F., Becker C. M. (2007) A novel glycine receptor β subunit splice variant predicts an unorthodox transmembrane topology. Assembly into heteromeric receptor complexes. J. Biol. Chem. 282, 2798–2807 - PubMed
-
- Harvey R. J., Topf M., Harvey K., Rees M. I. (2008) The genetics of hyperekplexia: More than startle! Trends Genet. 24, 439–447 - PubMed
-
- Kingsmore S. F., Giros B., Suh D., Bieniarz M., Caron M. G., Seldin M. F. (1994) Glycine receptor β-subunit gene mutation in spastic mouse associated with LINE-1 element insertion. Nat. Genet. 7, 136–141 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
