

Tmem100, an ALK1 receptor signaling-dependent gene essential for arterial endothelium differentiation and vascular morphogenesis
- PMID: 22783020
- PMCID: PMC3409742
- DOI: 10.1073/pnas.1207210109
Tmem100, an ALK1 receptor signaling-dependent gene essential for arterial endothelium differentiation and vascular morphogenesis
Abstract
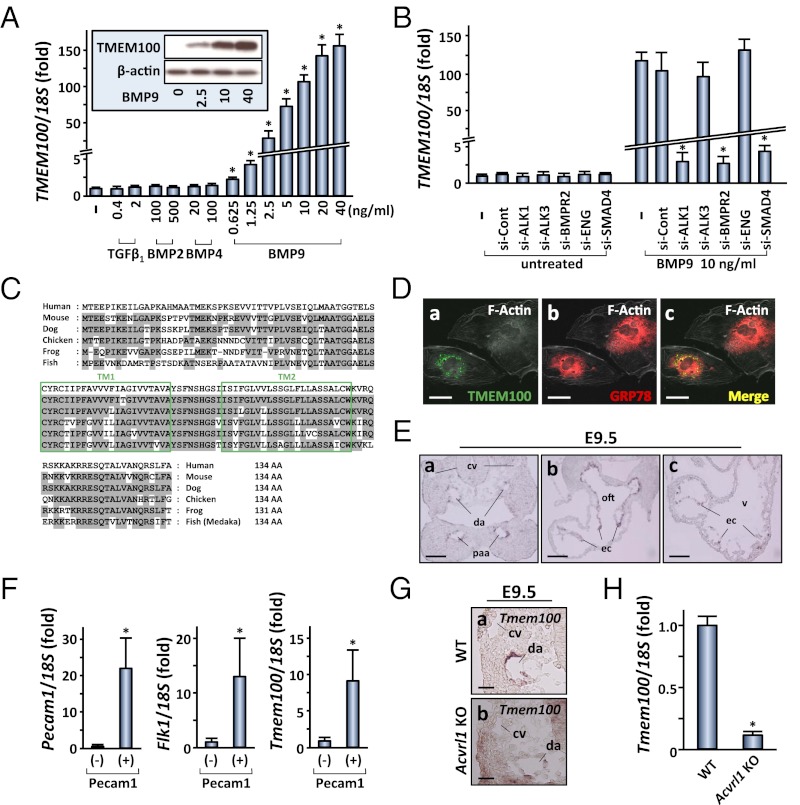
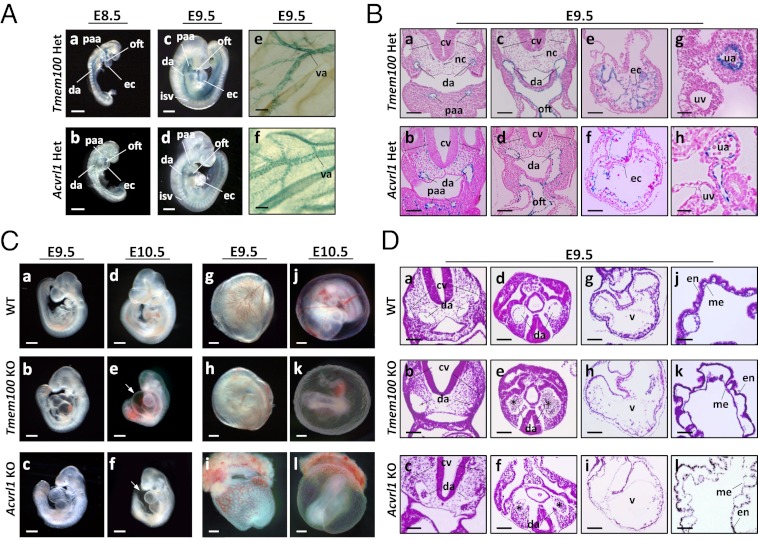
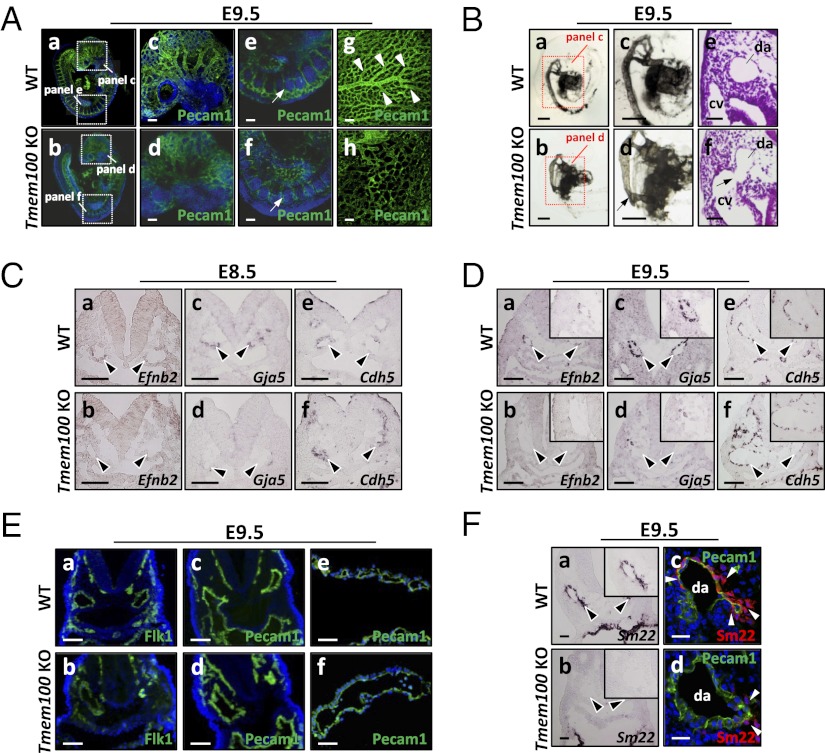
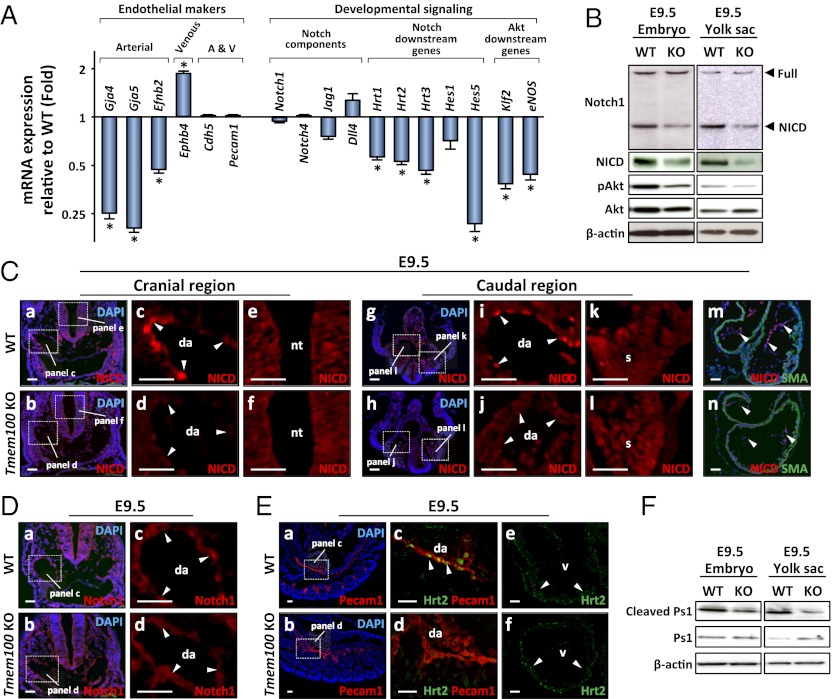
Members of the transforming growth factor-β superfamily play essential roles in various aspects of embryonic development and physiological organ function. Among them, bone morphogenetic protein (BMP) 9 and BMP10 regulate embryonic vascular development by activating their endothelial receptor ALK1 (activin receptor-like kinase 1, also called Acvrl1). ALK1-mediated intracellular signaling is implicated in the etiologies of human diseases, but their downstream functional proteins are largely unknown. In this study, we identified Tmem100, a gene encoding a previously uncharacterized intracellular transmembrane protein, to be an embryonic endothelium-enriched gene activated by BMP9 and BMP10 through the ALK1 receptor. Tmem100 null mice showed embryonic lethality due to impaired differentiation of arterial endothelium and defects of vascular morphogenesis, which phenocopied most of the vascular abnormalities observed with the Acvrl1/Alk1 deficiency. The activity of Notch- and Akt-mediated signaling, which is essential for vascular development, was down-regulated in Tmem100 null mice. Cre-mediated deletion of Tmem100 in endothelial cells was sufficient to recapitulate the null phenotypes. These data indicated that TMEM100 may play indispensable roles downstream of BMP9/BMP10-ALK1 signaling during endothelial differentiation and vascular morphogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Potente M, Gerhardt H, Carmeliet P. Basic and therapeutic aspects of angiogenesis. Cell. 2011;146:873–887. - PubMed
-
- Eilken HM, Adams RH. Dynamics of endothelial cell behavior in sprouting angiogenesis. Curr Opin Cell Biol. 2010;22:617–625. - PubMed
-
- David L, Feige JJ, Bailly S. Emerging role of bone morphogenetic proteins in angiogenesis. Cytokine Growth Factor Rev. 2009;20:203–212. - PubMed
-
- Pardali E, Goumans MJ, ten Dijke P. Signaling by members of the TGF-beta family in vascular morphogenesis and disease. Trends Cell Biol. 2010;20:556–567. - PubMed
-
- David L, Mallet C, Mazerbourg S, Feige JJ, Bailly S. Identification of BMP9 and BMP10 as functional activators of the orphan activin receptor-like kinase 1 (ALK1) in endothelial cells. Blood. 2007;109:1953–1961. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
