

Membrane protein structure determination using crystallography and lipidic mesophases: recent advances and successes
- PMID: 22783824
- PMCID: PMC3461499
- DOI: 10.1021/bi300010w
Membrane protein structure determination using crystallography and lipidic mesophases: recent advances and successes
Abstract
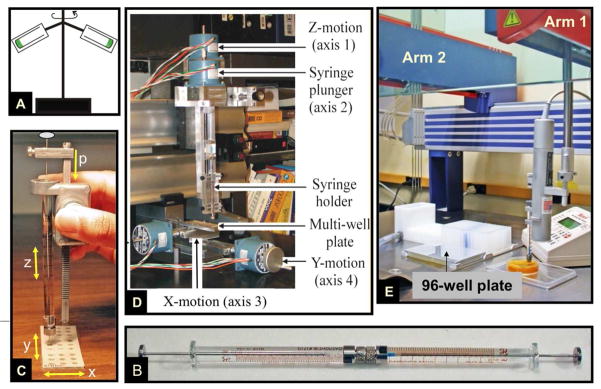
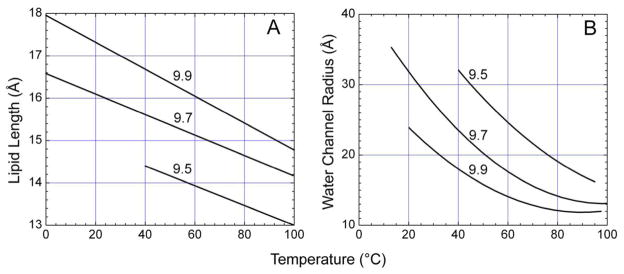
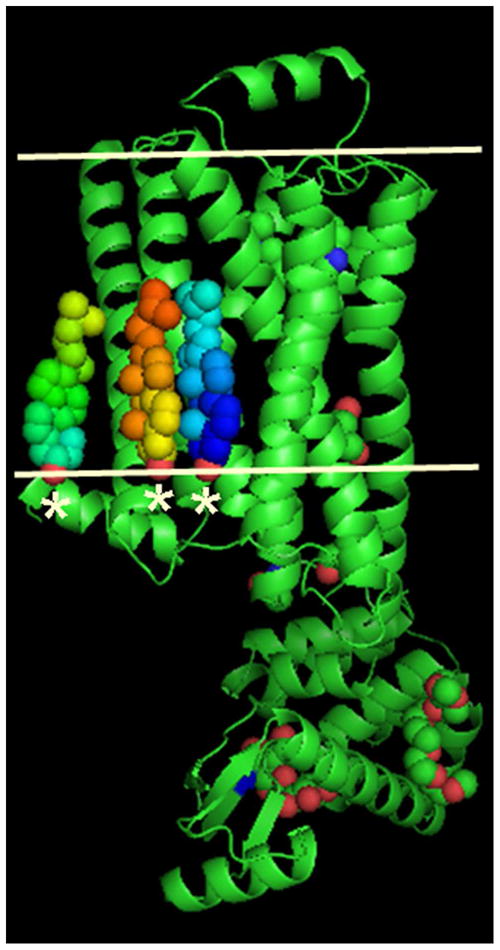
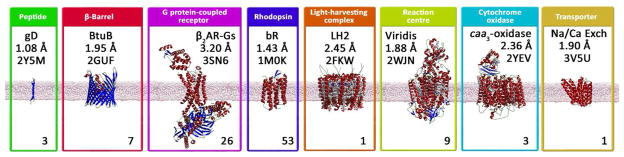
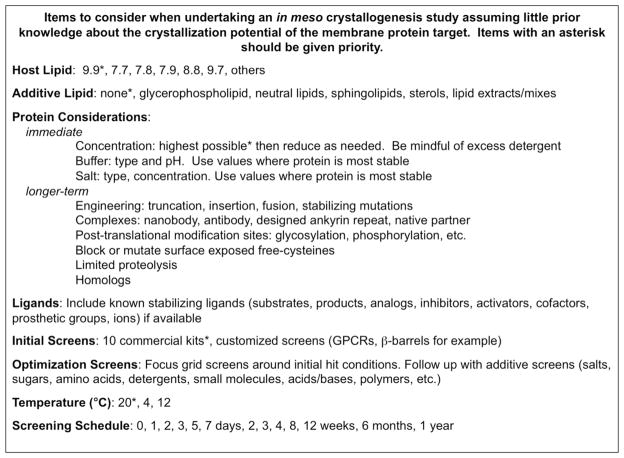
The crystal structure of the β(2)-adrenergic receptor in complex with an agonist and its cognate G protein has just recently been determined. It is now possible to explore in molecular detail the means by which this paradigmatic transmembrane receptor binds agonist, communicates the impulse or signaling event across the membrane, and sets in motion a series of G protein-directed intracellular responses. The structure was determined using crystals of the ternary complex grown in a rationally designed lipidic mesophase by the so-called in meso method. The method is proving to be particularly useful in the G protein-coupled receptor field where the structures of 13 distinct receptor types have been determined in the past 5 years. In addition to receptors, the method has proven to be useful with a wide variety of integral membrane protein classes that include bacterial and eukaryotic rhodopsins, light-harvesting complex II (LHII), photosynthetic reaction centers, cytochrome oxidases, β-barrels, an exchanger, and an integral membrane peptide. This attests to the versatility and range of the method and supports the view that the in meso method should be included in the arsenal of the serious membrane structural biologist. For this to happen, however, the reluctance to adopt it attributable, in part, to the anticipated difficulties associated with handling the sticky, viscous cubic mesophase in which crystals grow must be overcome. Harvesting and collecting diffraction data with the mesophase-grown crystals are also viewed with some trepidation. It is acknowledged that there are challenges associated with the method. Over the years, we have endeavored to establish how the method works at a molecular level and to make it user-friendly. To these ends, tools for handling the mesophase in the pico- to nanoliter volume range have been developed for highly efficient crystallization screening in manual and robotic modes. Methods have been implemented for evaluating the functional activity of membrane proteins reconstituted into the bilayer of the cubic phase as a prelude to crystallogenesis. Glass crystallization plates that provide unparalleled optical quality and sensitivity to nascent crystals have been built. Lipid and precipitant screens have been designed for a more rational approach to crystallogenesis such that the method can now be applied to an even wider variety of membrane protein types. In this work, these assorted advances are outlined along with a summary of the membrane proteins that have yielded to the method. The prospects for and the challenges that must be overcome to further develop the method are described.
Figures
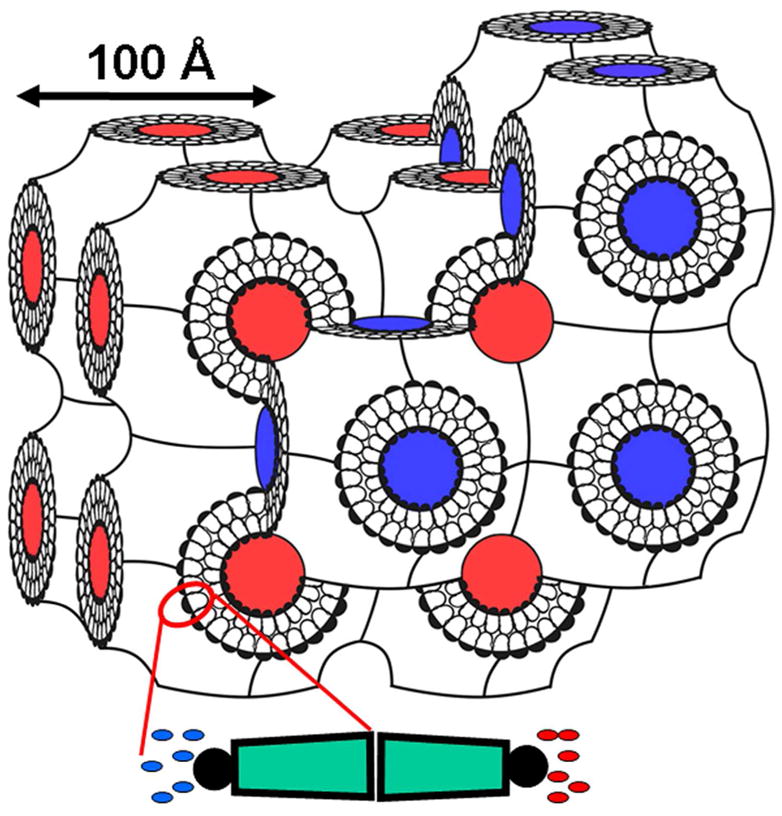
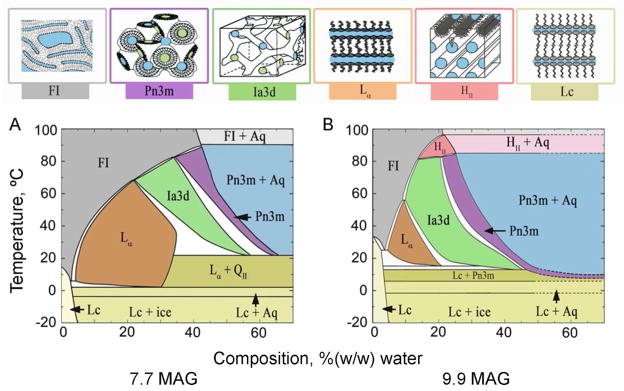



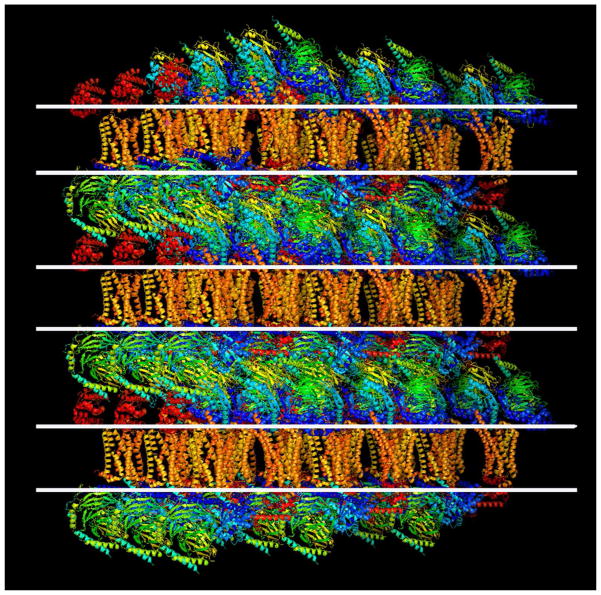
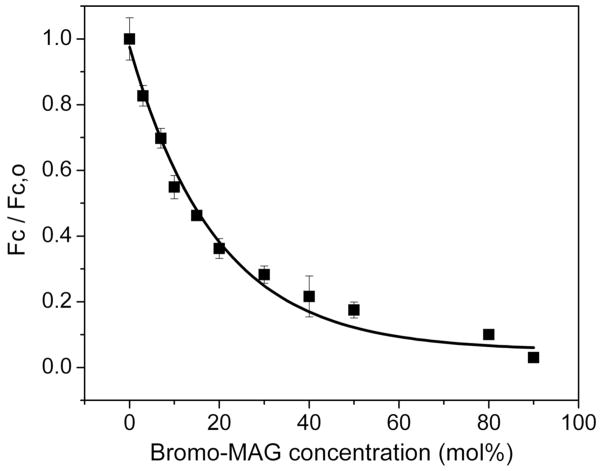
References
-
- Caffrey M. Membrane protein crystallization. J Struct Biol. 2003;142:108–132. - PubMed
-
- Michel H. Crystallization of membrane-proteins. Trends Biochem Sci. 1983;8:56–59.
-
- Wada T, Shimono K, Kikukawa T, Hato M, Shinya N, Kim SY, Kimura-Someya T, Shirouzu M, Tamogami J, Miyauchi S, Jung KH, Kamo N, Yokoyama S. Crystal structure of the eukaryotic light-driven proton-pumping rhodopsin, Acetabularia rhodopsin II, from marine alga. J Mol Biol. 2011;411:986–998. - PubMed
-
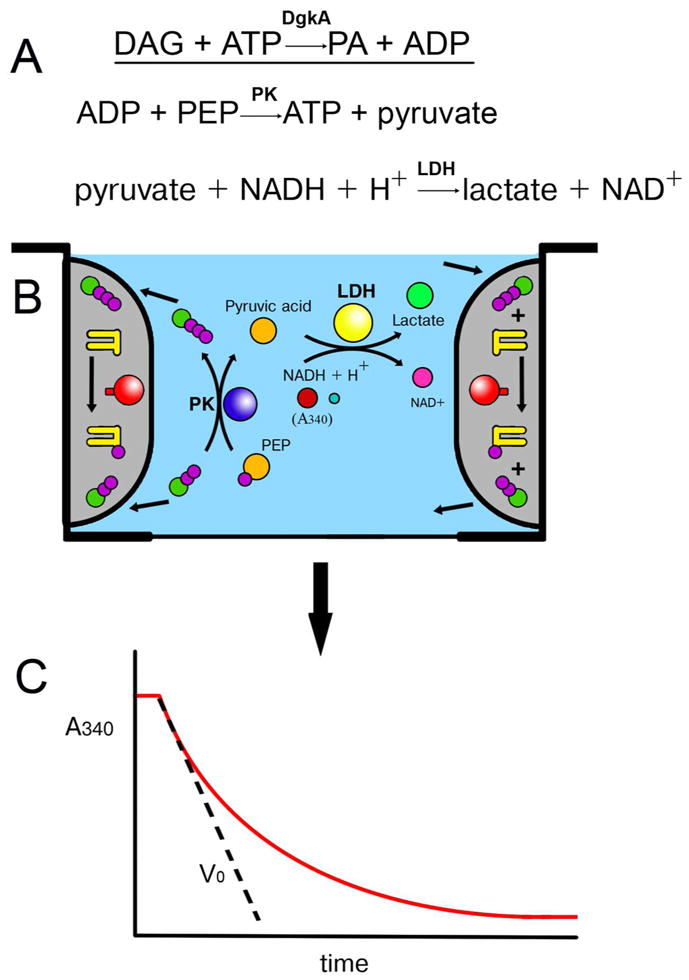
- Lahiri S, Brehs M, Olschewski D, Becker CFW. Total chemical synthesis of an integral membrane enzyme: Diacylglycerol kinase from Escherichia coli. Angewandte Chemie-International Edition. 2011;50:3988–3992. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
