

Induction of robust de novo centrosome amplification, high-grade spindle multipolarity and metaphase catastrophe: a novel chemotherapeutic approach
- PMID: 22785532
- PMCID: PMC3406581
- DOI: 10.1038/cddis.2012.82
Induction of robust de novo centrosome amplification, high-grade spindle multipolarity and metaphase catastrophe: a novel chemotherapeutic approach
Abstract
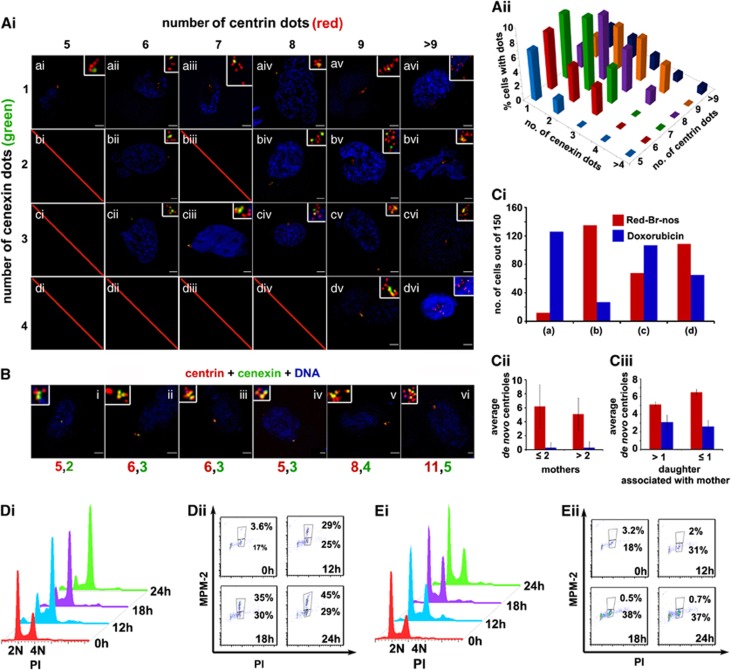
Centrosome amplification (CA) and resultant chromosomal instability have long been associated with tumorigenesis. However, exacerbation of CA and relentless centrosome declustering engender robust spindle multipolarity (SM) during mitosis and may induce cell death. Recently, we demonstrated that a noscapinoid member, reduced bromonoscapine, (S)-3-(R)-9-bromo-5-(4,5-dimethoxy-1,3-dihydroisobenzofuran-1-yl)-4-methoxy-6-methyl-5,6,7,8-tetrahydro-[1,3]dioxolo-[4,5-g]isoquinoline (Red-Br-nos), induces reactive oxygen species (ROS)-mediated autophagy and caspase-independent death in prostate cancer PC-3 cells. Herein, we show that Red-Br-nos induces ROS-dependent DNA damage that resulted in high-grade CA and SM in PC-3 cells. Unlike doxorubicin, which causes double-stranded DNA breaks and chronic G2 arrest accompanied by 'templated' CA, Red-Br-nos-mediated DNA damage elicits de novo CA during a transient S/G2 stall, followed by checkpoint abrogation and mitotic entry to form aberrant mitotic figures with supernumerary spindle poles. Attenuation of multipolar phenotype in the presence of tiron, a ROS inhibitor, indicated that ROS-mediated DNA damage was partly responsible for driving CA and SM. Although a few cells (∼5%) yielded to aberrant cytokinesis following an 'anaphase catastrophe', most mitotically arrested cells (∼70%) succumbed to 'metaphase catastrophe,' which was caspase-independent. This report is the first documentation of rapid de novo centrosome formation in the presence of parent centrosome by a noscapinoid family member, which triggers death-inducing SM via a unique mechanism that distinguishes it from other ROS-inducers, conventional DNA-damaging agents, as well as other microtubule-binding drugs.
Figures






References
-
- Lingle WL, Lukasiewicz K, Salisbury JL. Deregulation of the centrosome cycle and the origin of chromosomal instability in cancer. Adv Exp Med Biol. 2005;570:393–421. - PubMed
-
- Brinkley BR. Managing the centrosome numbers game: from chaos to stability in cancer cell division. Trends Cell Biol. 2001;11:18–21. - PubMed
-
- Quintyne NJ, Reing JE, Hoffelder DR, Gollin SM, Saunders WS. Spindle multipolarity is prevented by centrosomal clustering. Science. 2005;307:127–129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
