

Song tutoring in presinging zebra finch juveniles biases a small population of higher-order song-selective neurons toward the tutor song
- PMID: 22786956
- PMCID: PMC3544995
- DOI: 10.1152/jn.00905.2011
Song tutoring in presinging zebra finch juveniles biases a small population of higher-order song-selective neurons toward the tutor song
Abstract
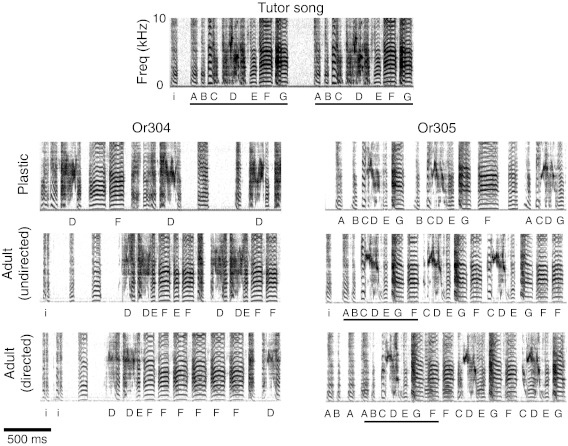
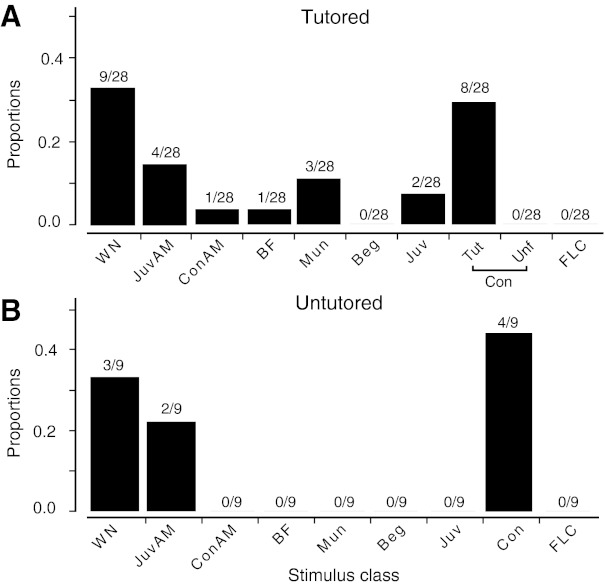
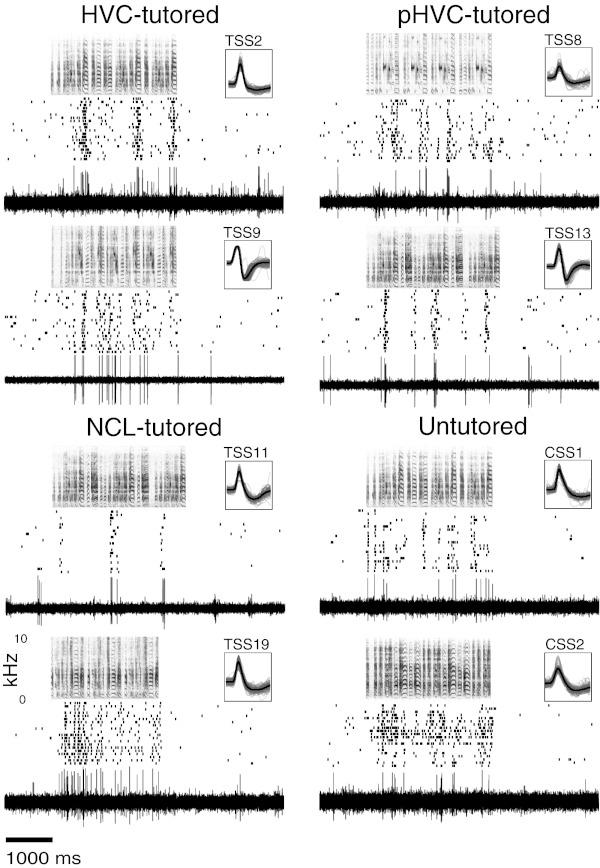
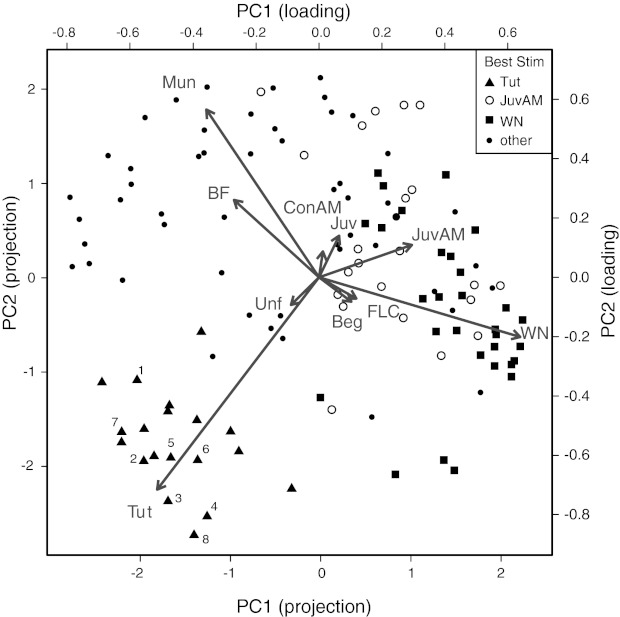
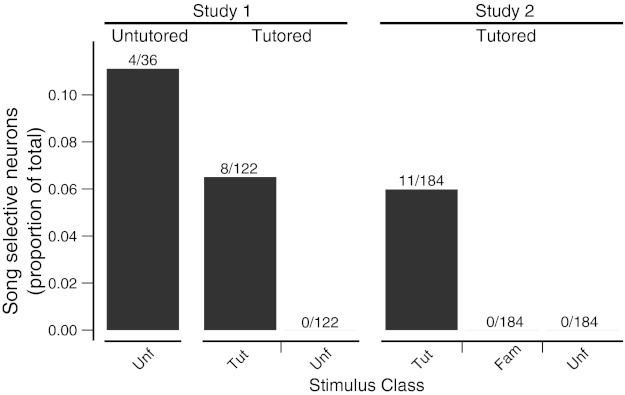
We explored physiological changes correlated with song tutoring by recording the responses of caudal nidopallium neurons of zebra finches aged P21-P24 (days post hatching) to a broad spectrum of natural and synthetic stimuli. Those birds raised with their fathers tended to show behavioral evidence of song memorization but not of singing; thus auditory responses were not confounded by the birds' own vocalizations. In study 1, 37 of 158 neurons (23%) in 17 of 22 tutored and untutored birds were selective for only 1 of 10 stimuli comprising broadband signals, early juvenile songs and calls, female calls, and adult songs. Approximately 30% of the selective neurons (12/37 neurons in 9 birds) were selective for adult conspecific songs. All these were found in the song system nuclei HVC and paraHVC. Of 122 neurons (17 birds) in tutored birds, all of the conspecific song-selective neurons (8 neurons in 6 birds) were selective for the adult tutor song; none was selective for unfamiliar song. In study 2 with a different sampling strategy, we found that 11 of 12 song-selective neurons in 6 of 7 birds preferred the tutor song; none preferred unfamiliar or familiar conspecific songs. Most of these neurons were found in caudal lateral nidopallium (NCL) below HVC. Thus by the time a bird begins to sing, there are small numbers of tutor song-selective neurons distributed in several forebrain regions. We hypothesize that a small population of higher-order auditory neurons is innately selective for complex features of behaviorally relevant stimuli and these responses are modified by specific perceptual/social experience during development.
Figures
References
-
- Adret P. Operant conditioning, song learning and imprinting to taped song in the zebra finch. Anim Behav 45: 149–159, 1993
-
- Adret P. The template concept: crafting a song replica from memory. In: Neuroscience of Birdsong, edited by Zeigler HP, Marler P. Cambridge, UK: Cambridge Univ. Press, 2008, p. 282–299
-
- Aronov D, Andalman AS, Fee MS. A specialized forebrain circuit for vocal babbling in the juvenile songbird. Science 320: 630–634, 2008 - PubMed
-
- Baptista LF, Petrinovich L. Social interaction, sensitive phases and the song template hypothesis in the white-crowned sparrow. Anim Behav 32: 172–181, 1984
-
- Basham ME, Nordeen EJ, Nordeen KW. Blockade of NMDA receptors in the anterior forebrain impairs sensory acquisition in the zebra finch (Poephila guttata). Neurobiol Learn Mem 66: 295–304, 1996 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous