

Herpes simplex virus is equipped with RNA- and protein-based mechanisms to repress expression of ATRX, an effector of intrinsic immunity
- PMID: 22787211
- PMCID: PMC3446562
- DOI: 10.1128/JVI.00930-12
Herpes simplex virus is equipped with RNA- and protein-based mechanisms to repress expression of ATRX, an effector of intrinsic immunity
Abstract
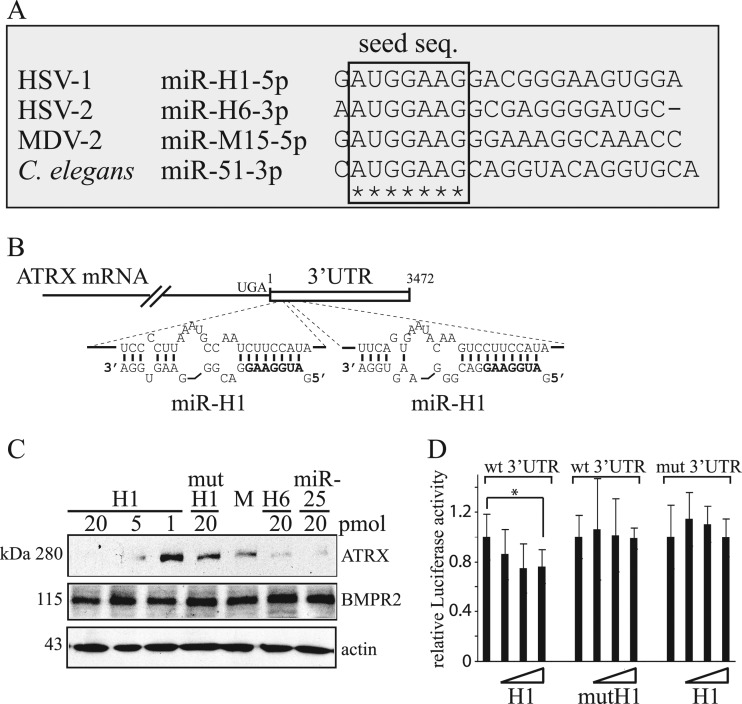
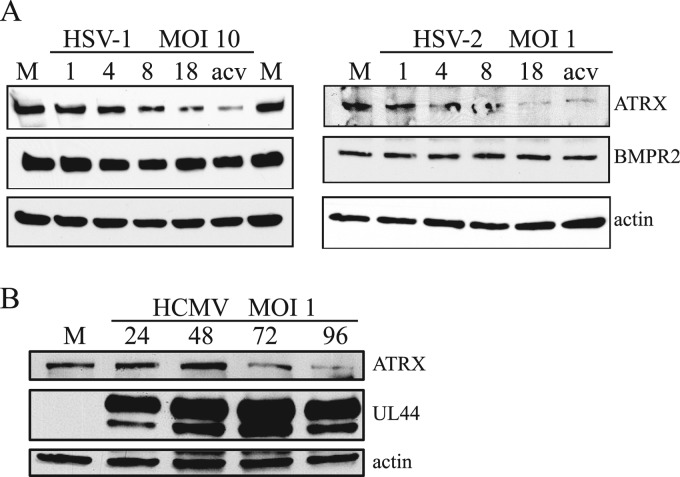
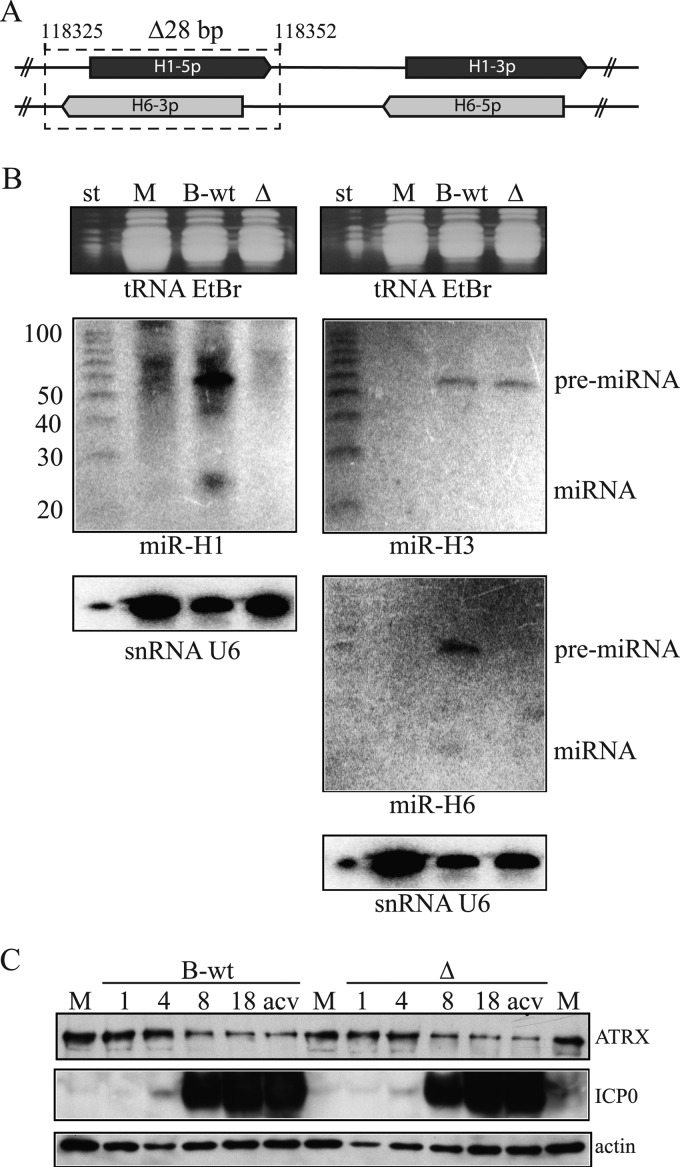
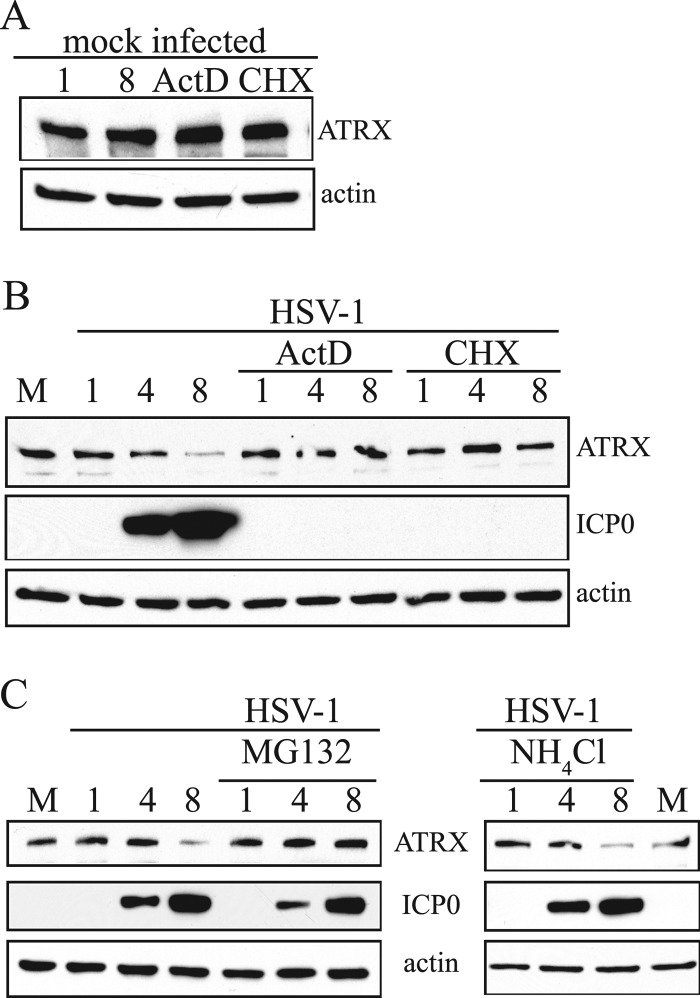
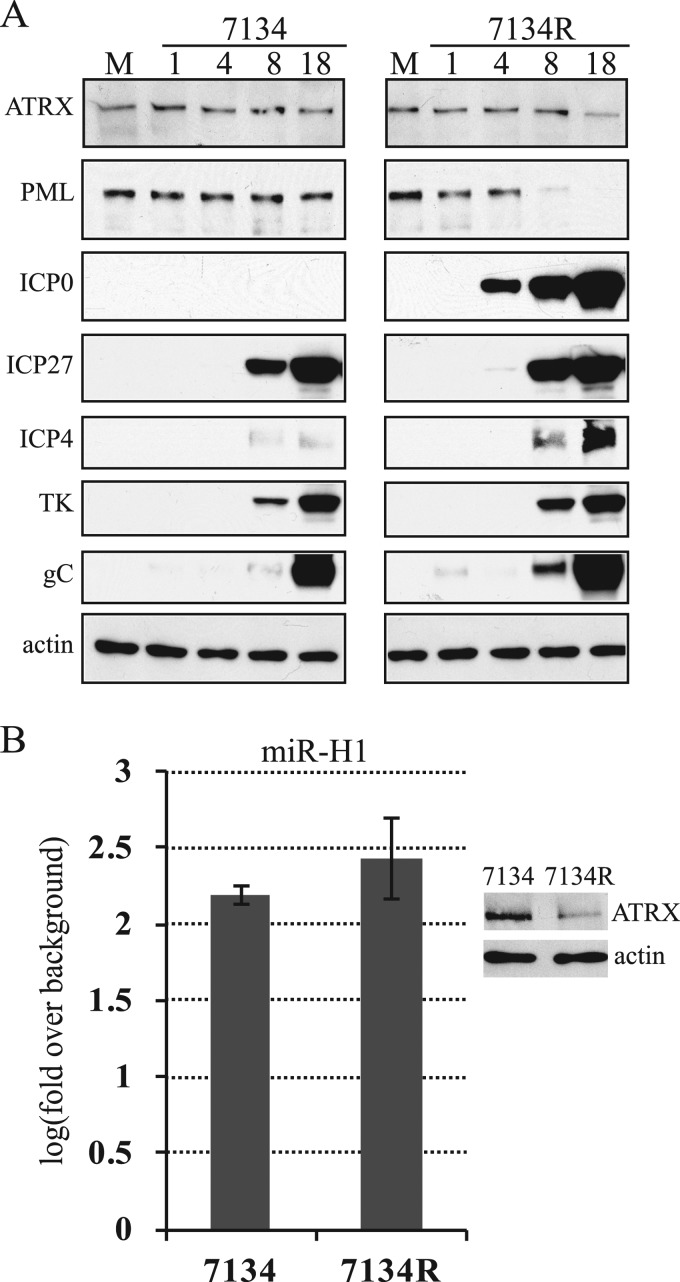
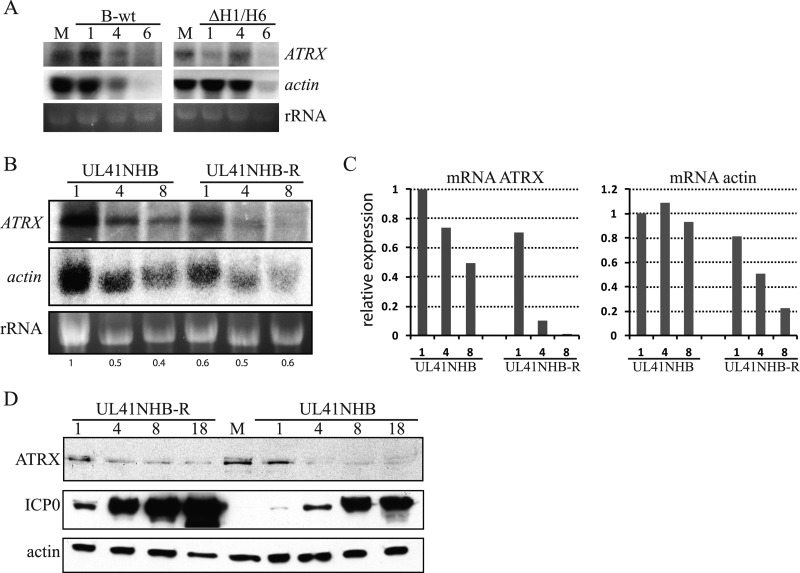
Intrinsic immunity is a first-line intracellular defense against virus infection, and viruses have evolved mechanisms to counteract it. During herpes simplex virus (HSV) infection, nuclear domain 10 (ND10) components localize adjacent to incoming viral genomes and generate a repressive environment for viral gene expression. Here, we found that the ND10 component, alpha-thalassemia/mental retardation syndrome X-linked (ATRX) protein, is predicted to be a target of HSV-1 miR-H1 and HSV-2 miR-H6. These microRNAs (miRNAs) share a seed sequence and are abundant during lytic infection. Mimics of both miRNAs could deplete endogenous ATRX, and an miR-H1 mimic could repress the expression of a reporter linked to the 3' untranslated region of ATRX mRNA, identifying a cellular mRNA targeted by an HSV miRNA. Interestingly, ATRX protein and its mRNA were depleted in cells lytically infected with HSV, and ATRX protein was also depleted in cells infected with human cytomegalovirus. However, infection with an HSV-1 mutant lacking miR-H1 still resulted in ATRX depletion. This depletion was sensitive to a proteasome inhibitor and was largely ablated by a deletion of the gene encoding the immediate-early ICP0 protein. Additionally, a deletion of the gene encoding the tegument protein Vhs ablated most of the depletion of ATRX mRNA. Thus, HSV is equipped with multiple mechanisms to limit the expression of ATRX. As ATRX is implicated in repression of lytic viral gene expression, our results suggest roles for these different mechanisms during various phases of HSV infection.
Figures
References
-
- Adler M, Tavalai N, Muller R, Stamminger T. 2011. Human cytomegalovirus immediate-early gene expression is restricted by the nuclear domain 10 component Sp100. J. Gen. Virol. 92:1532–1538 - PubMed
-
- Becker Y, Tavor E, Asher Y, Berkowitz C, Moyal M. 1993. Effect of herpes simplex virus type-1 UL41 gene on the stability of mRNA from the cellular genes: beta-actin, fibronectin, glucose transporter-1, and docking protein, and on virus intraperitoneal pathogenicity to newborn mice. Virus Genes 7:133–143 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
