

ORF45 of Kaposi's sarcoma-associated herpesvirus inhibits phosphorylation of interferon regulatory factor 7 by IKKε and TBK1 as an alternative substrate
- PMID: 22787218
- PMCID: PMC3446610
- DOI: 10.1128/JVI.05224-11
ORF45 of Kaposi's sarcoma-associated herpesvirus inhibits phosphorylation of interferon regulatory factor 7 by IKKε and TBK1 as an alternative substrate
Abstract
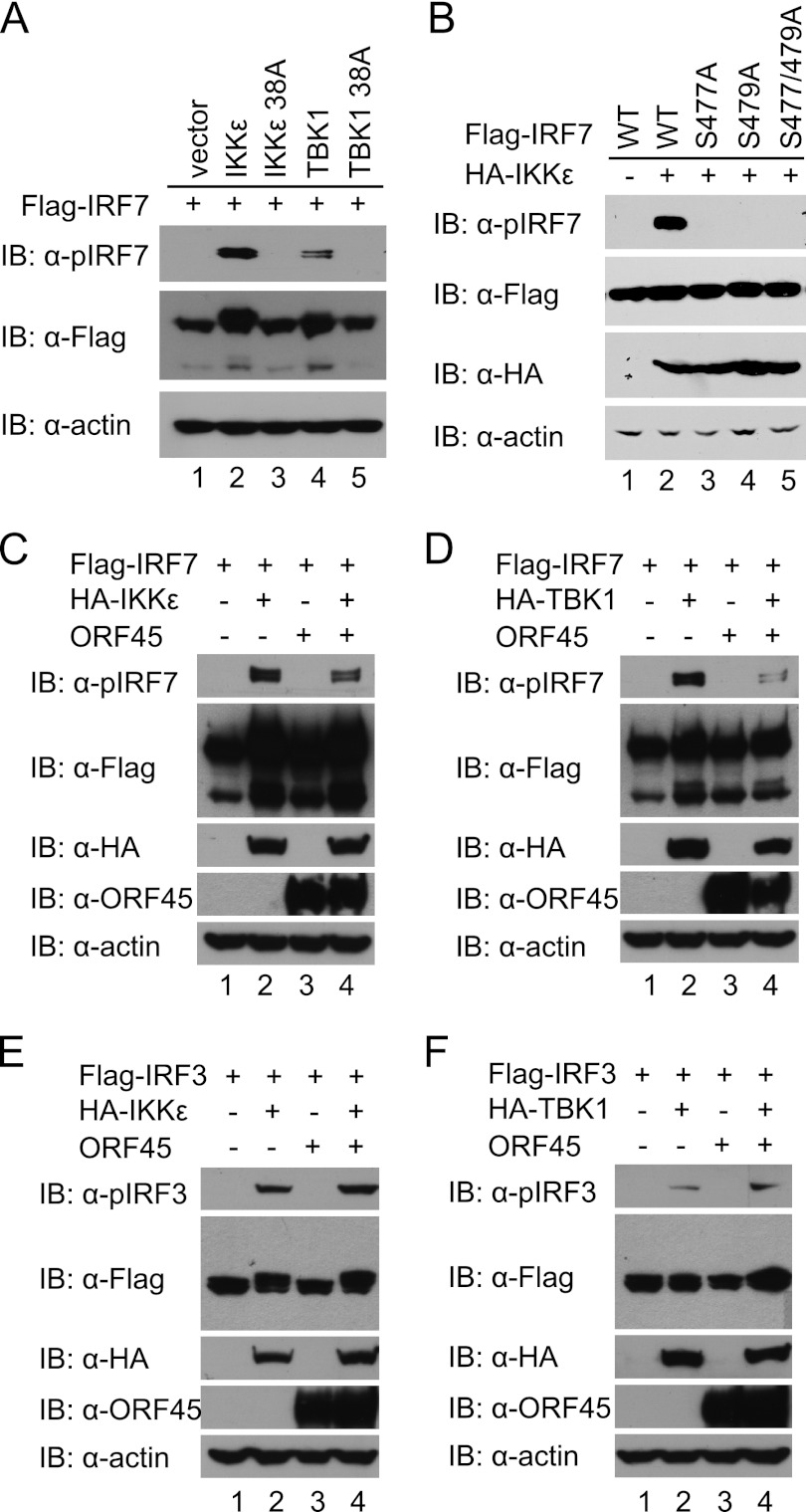
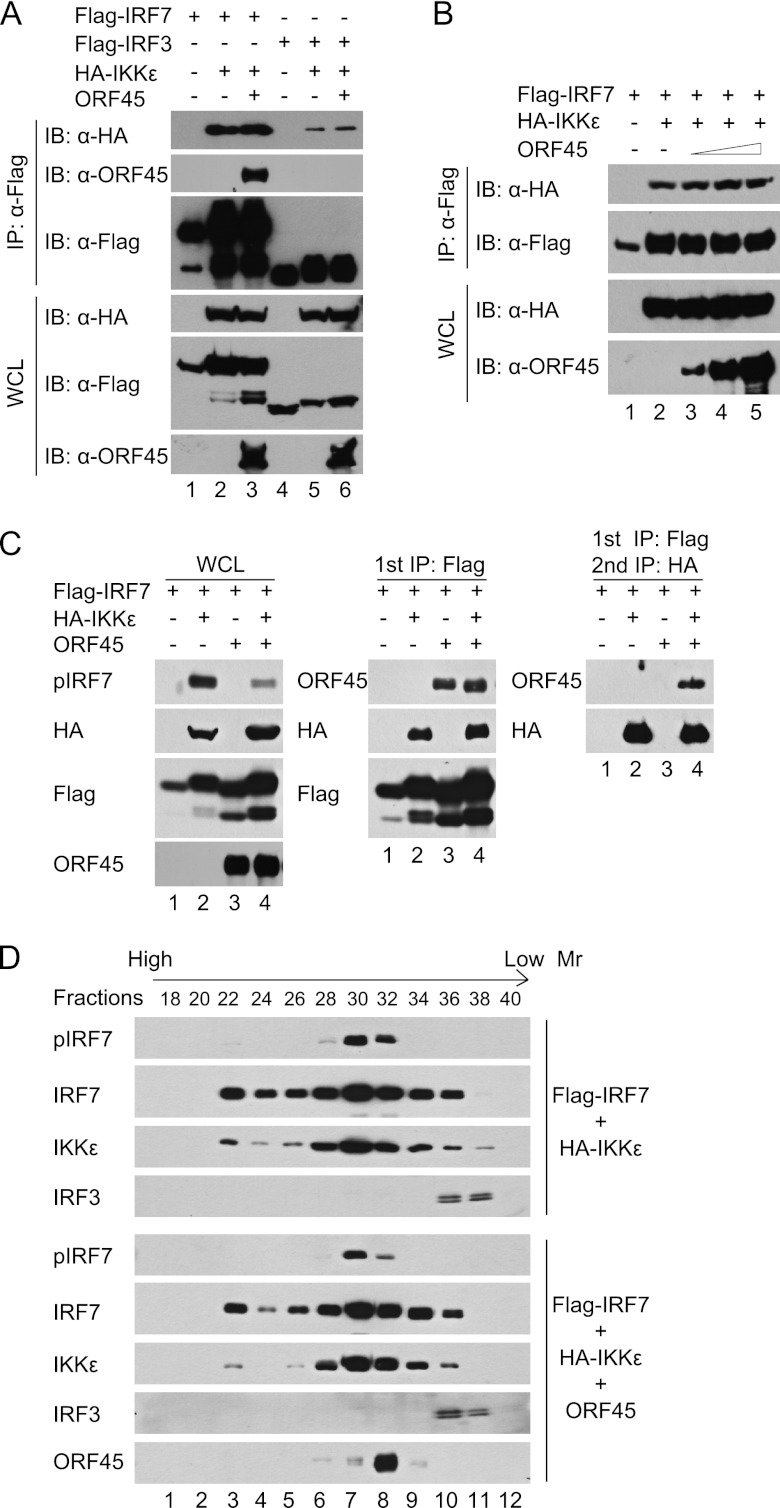
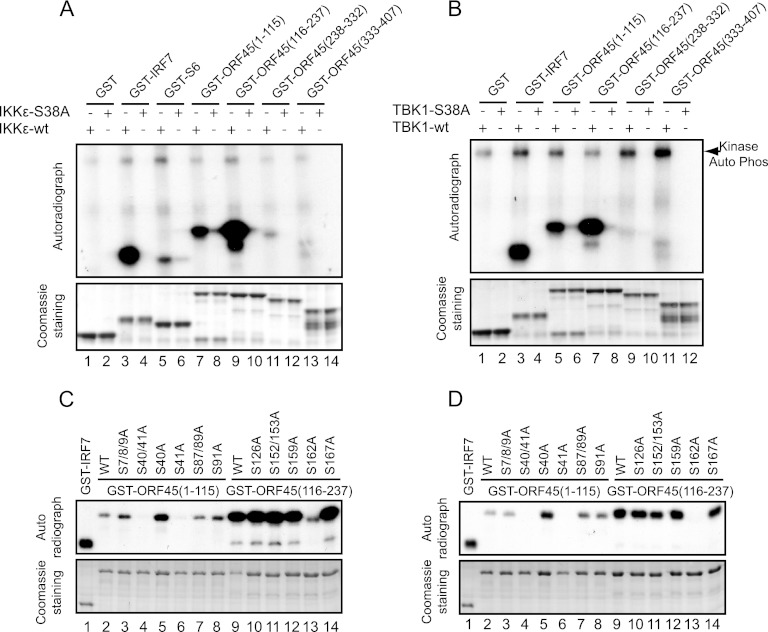
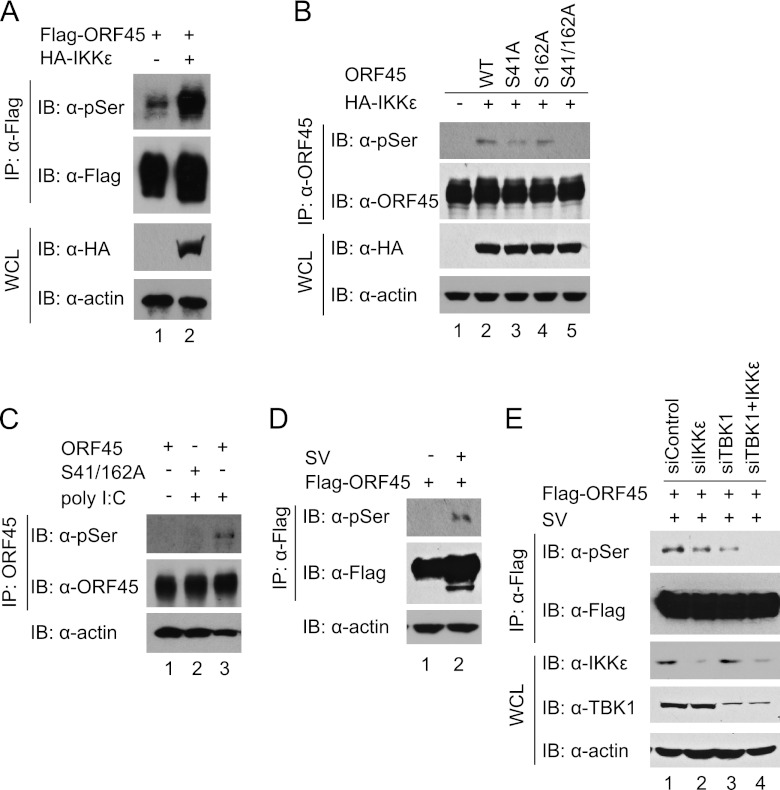
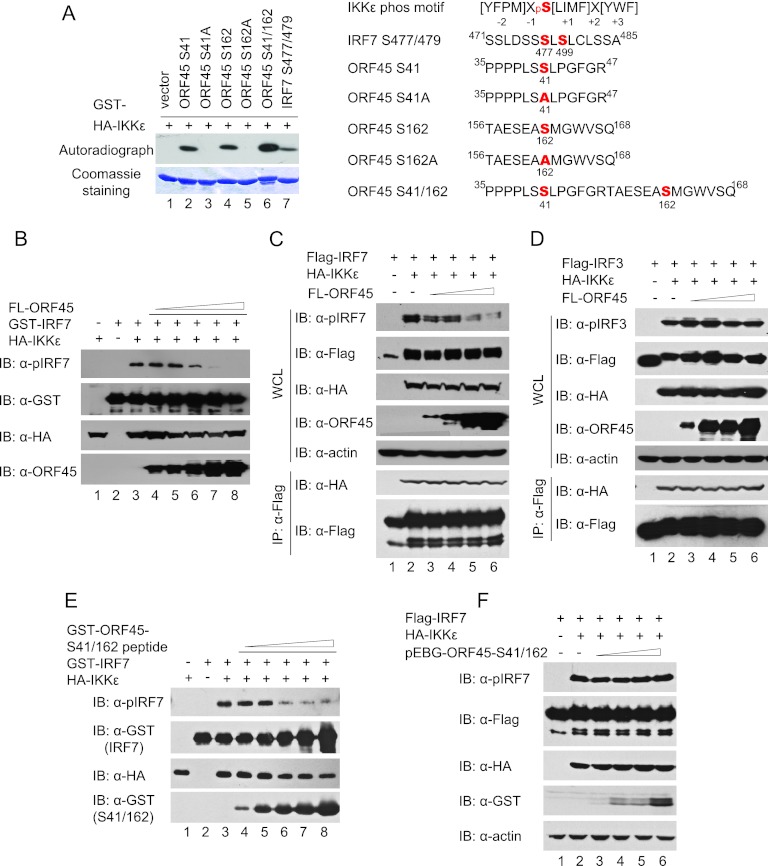
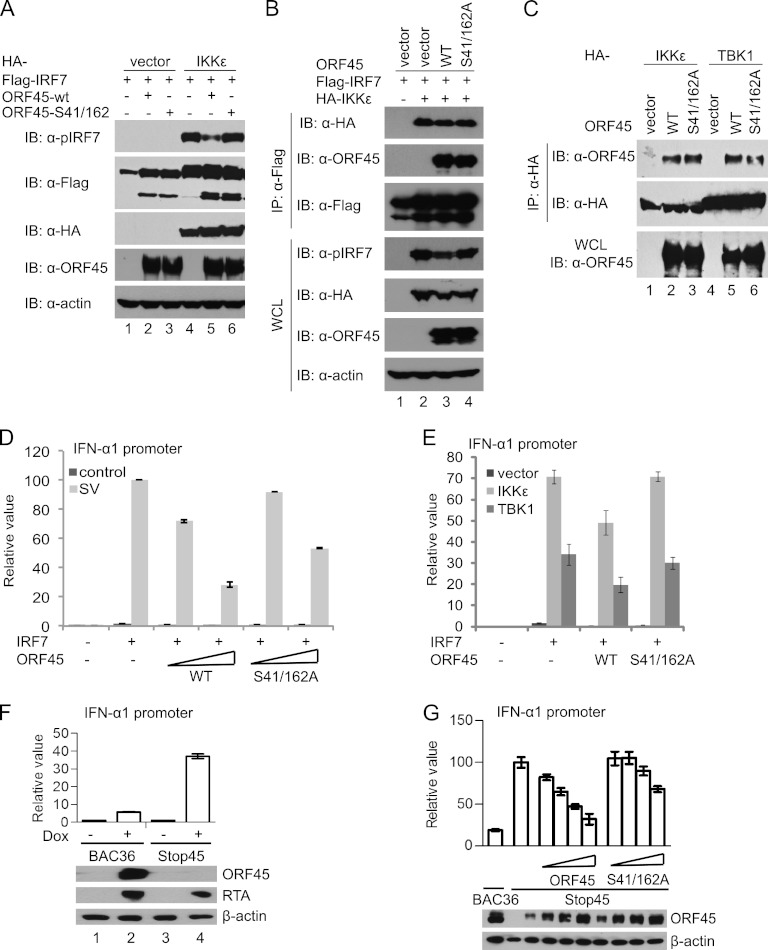
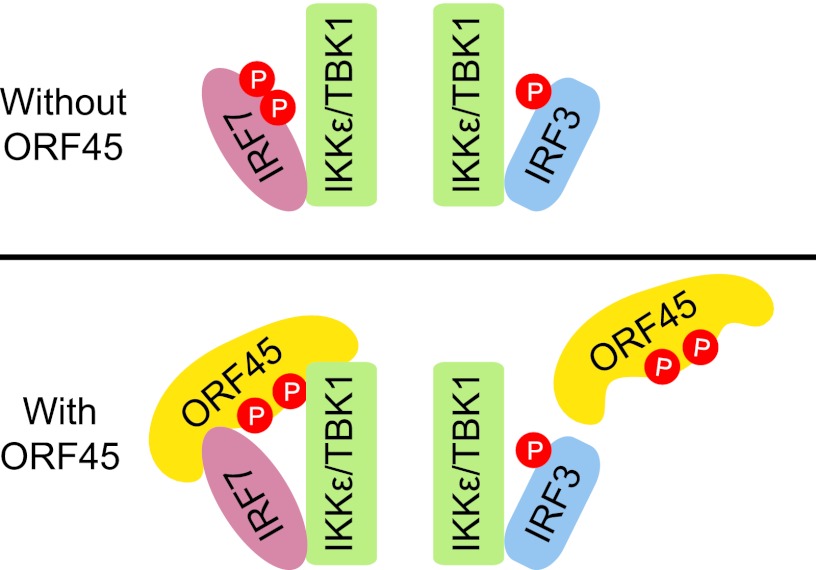
Open reading frame 45 (ORF45) of Kaposi's sarcoma-associated herpesvirus (KSHV) is an immediate-early and tegument protein that plays critical roles in antagonizing host antiviral responses. We have previously shown (Zhu et al, Proc. Natl. Acad. Sci. U. S. A., 99:5573-5578, 2002) that ORF45 suppresses activation of interferon regulatory factor 7 (IRF7), a crucial regulator of type I interferon gene expression, by blocking its virus-induced phosphorylation and nuclear accumulation. We report here further characterization of the mechanisms by which ORF45 inhibits IRF7 phosphorylation. In most cell types, IRF7 is phosphorylated and activated by IKKε and TBK1 after viral infection. We found that phosphorylation of IRF7 on Ser477 and Ser479 by IKKε or TBK1 is inhibited by ORF45. The inhibition is specific to IRF7 because phosphorylation of its close relative IRF3 is not affected by ORF45, implying that ORF45 does not inactivate the kinases directly. In fact, we found that ORF45 is phosphorylated efficiently on Ser41 and Ser162 by IKKε and TBK1. We demonstrated that ORF45 competes with the associated IRF7 and inhibits its phosphorylation by IKKε or TBK1 by acting as an alternative substrate.
Figures
Similar articles
-
The ORF45 Protein of Kaposi's Sarcoma-Associated Herpesvirus and Its Critical Role in the Viral Life Cycle.Viruses. 2022 Sep 11;14(9):2010. doi: 10.3390/v14092010. Viruses. 2022. PMID: 36146816 Free PMC article. Review.
-
The ORF45 protein of Kaposi's sarcoma-associated herpesvirus is associated with purified virions.J Virol. 2003 Apr;77(7):4221-30. doi: 10.1128/jvi.77.7.4221-4230.2003. J Virol. 2003. PMID: 12634379 Free PMC article.
-
Mechanisms of autoinhibition of IRF-7 and a probable model for inactivation of IRF-7 by Kaposi's sarcoma-associated herpesvirus protein ORF45.J Biol Chem. 2011 Jan 7;286(1):746-56. doi: 10.1074/jbc.M110.150920. Epub 2010 Oct 27. J Biol Chem. 2011. PMID: 20980251 Free PMC article.
-
Antagonism of host antiviral responses by Kaposi's sarcoma-associated herpesvirus tegument protein ORF45.PLoS One. 2010 May 11;5(5):e10573. doi: 10.1371/journal.pone.0010573. PLoS One. 2010. PMID: 20485504 Free PMC article.
-
ORF45, a multifunctional immediate early and tegument protein of KSHV.J Med Virol. 2023 Mar;95(3):e28659. doi: 10.1002/jmv.28659. J Med Virol. 2023. PMID: 36905218 Review.
Cited by
-
The ORF45 Protein of Kaposi's Sarcoma-Associated Herpesvirus and Its Critical Role in the Viral Life Cycle.Viruses. 2022 Sep 11;14(9):2010. doi: 10.3390/v14092010. Viruses. 2022. PMID: 36146816 Free PMC article. Review.
-
The SUMO E3 ligase activity of ORF45 determines KSHV lytic replication.PLoS Pathog. 2022 Apr 28;18(4):e1010504. doi: 10.1371/journal.ppat.1010504. eCollection 2022 Apr. PLoS Pathog. 2022. PMID: 35482828 Free PMC article.
-
KSHV hijacks the antiviral kinase IKKε to initiate lytic replication.PLoS Pathog. 2025 Jan 17;21(1):e1012856. doi: 10.1371/journal.ppat.1012856. eCollection 2025 Jan. PLoS Pathog. 2025. PMID: 39823515 Free PMC article.
-
Evasion of adaptive and innate immune response mechanisms by γ-herpesviruses.Curr Opin Virol. 2013 Jun;3(3):285-95. doi: 10.1016/j.coviro.2013.05.011. Epub 2013 Jun 2. Curr Opin Virol. 2013. PMID: 23735334 Free PMC article. Review.
-
ORF48 is required for optimal lytic replication of Kaposi's sarcoma-associated herpesvirus.PLoS Pathog. 2024 Aug 26;20(8):e1012081. doi: 10.1371/journal.ppat.1012081. eCollection 2024 Aug. PLoS Pathog. 2024. PMID: 39186813 Free PMC article.
References
-
- Adli M, Baldwin AS. 2006. IKK-i/IKKepsilon controls constitutive, cancer cell-associated NF-kappaB activity via regulation of Ser-536 p65/RelA phosphorylation. J. Biol. Chem. 281:26976–26984 - PubMed
-
- Boehm JS, et al. 2007. Integrative genomic approaches identify IKBKE as a breast cancer oncogene. Cell 129:1065–1079 - PubMed
-
- Buettner N, et al. 2010. Thogoto virus ML protein is a potent inhibitor of the interferon regulatory factor-7 transcription factor. J. Gen. Virol. 91:220–227 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
