

Base excision repair: contribution to tumorigenesis and target in anticancer treatment paradigms
- PMID: 22788768
- PMCID: PMC4537170
- DOI: 10.2174/092986712802002581
Base excision repair: contribution to tumorigenesis and target in anticancer treatment paradigms
Abstract
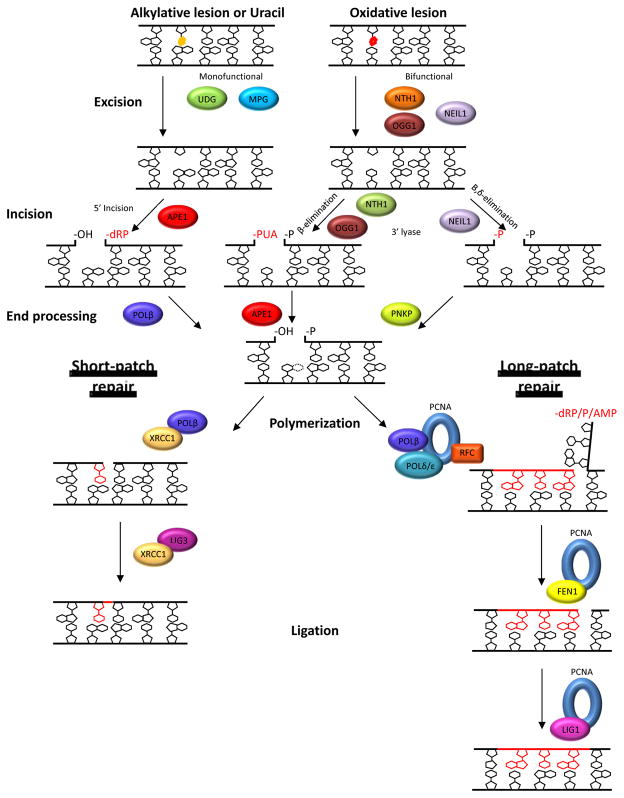
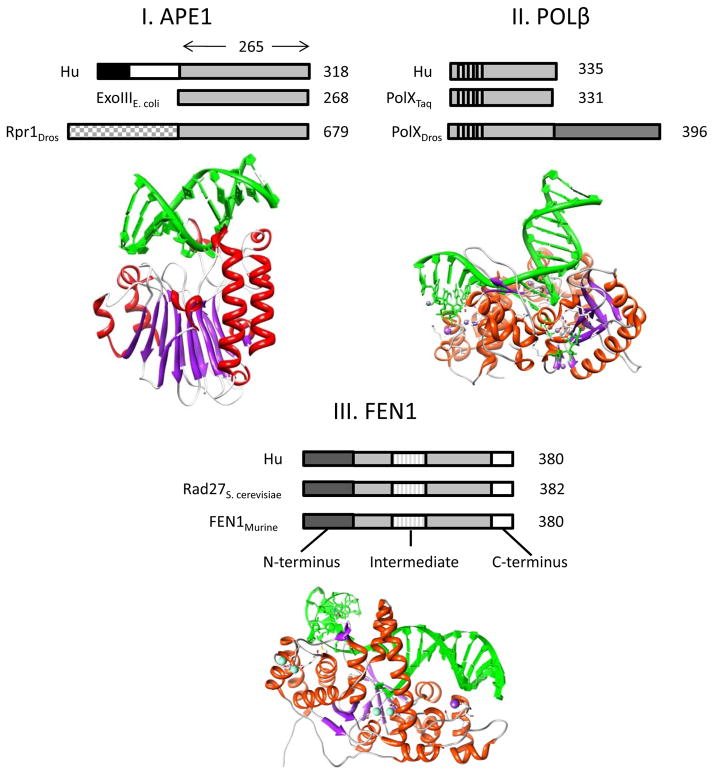
Cancer treatments often lose their effectiveness due to the development of multiple drug resistance. Thus, identification of key proteins involved in the tumorigenic process and the survival mechanism(s), coupled with the design of novel therapeutic compounds (such as small molecule inhibitors), are essential steps towards the establishment of improved anticancer treatment strategies. DNA repair pathways and their proteins have been exposed as potential targets for combinatorial anticancer therapies that involve DNA-interactive cytotoxins, such as alkylating agents, because of their central role in providing resistance against DNA damage. In addition, an understanding of the tumor-specific genetics and associated DNA repair capacity has allowed research scientists and clinicians to begin to devise more targeted treatment strategies based on the concept of synthetic lethality. In this review, the repair mechanisms, as well as the links to cancer progression and treatment, of three key proteins that function in the base excision repair pathway, i.e. APE1, POLβ, and FEN1, are discussed.
Figures
Similar articles
-
Human AP endonuclease 1 (APE1): from mechanistic insights to druggable target in cancer.Cancer Treat Rev. 2010 Aug;36(5):425-35. doi: 10.1016/j.ctrv.2009.12.006. Epub 2010 Jan 6. Cancer Treat Rev. 2010. PMID: 20056333 Review.
-
Study of interaction of XRCC1 with DNA and proteins of base excision repair by photoaffinity labeling technique.Biochemistry (Mosc). 2007 Aug;72(8):878-86. doi: 10.1134/s000629790708010x. Biochemistry (Mosc). 2007. PMID: 17922646
-
Single-molecule studies of repair proteins in base excision repair.BMB Rep. 2025 Jan;58(1):17-23. doi: 10.5483/BMBRep.2024-0178. BMB Rep. 2025. PMID: 39701025 Free PMC article. Review.
-
Inhibition of the human apurinic/apyrimidinic endonuclease (APE1) repair activity and sensitization of breast cancer cells to DNA alkylating agents with lucanthone.Anticancer Res. 2004 Jul-Aug;24(4):2127-34. Anticancer Res. 2004. PMID: 15330152
-
Inhibitors of nuclease and redox activity of apurinic/apyrimidinic endonuclease 1/redox effector factor 1 (APE1/Ref-1).Bioorg Med Chem. 2017 May 1;25(9):2531-2544. doi: 10.1016/j.bmc.2017.01.028. Epub 2017 Jan 21. Bioorg Med Chem. 2017. PMID: 28161249 Review.
Cited by
-
YU238259 Is a Novel Inhibitor of Homology-Dependent DNA Repair That Exhibits Synthetic Lethality and Radiosensitization in Repair-Deficient Tumors.Mol Cancer Res. 2015 Oct;13(10):1389-97. doi: 10.1158/1541-7786.MCR-15-0036. Epub 2015 Jun 26. Mol Cancer Res. 2015. PMID: 26116172 Free PMC article.
-
Identification of Novel Pathways Regulated by APE1/Ref-1 in Human Retinal Endothelial Cells.Int J Mol Sci. 2023 Jan 6;24(2):1101. doi: 10.3390/ijms24021101. Int J Mol Sci. 2023. PMID: 36674619 Free PMC article.
-
Repair of oxidatively induced DNA damage by DNA glycosylases: Mechanisms of action, substrate specificities and excision kinetics.Mutat Res Rev Mutat Res. 2017 Jan-Mar;771:99-127. doi: 10.1016/j.mrrev.2017.02.001. Epub 2017 Feb 16. Mutat Res Rev Mutat Res. 2017. PMID: 28342455 Free PMC article. Review.
-
FEN1 is a prognostic biomarker for ER+ breast cancer and associated with tamoxifen resistance through the ERα/cyclin D1/Rb axis.Ann Transl Med. 2021 Feb;9(3):258. doi: 10.21037/atm-20-3068. Ann Transl Med. 2021. PMID: 33708885 Free PMC article.
-
Polymorphic mutations in the polb gene promoter and their impact on transcriptional activity.Thorac Cancer. 2022 Mar;13(6):853-857. doi: 10.1111/1759-7714.14337. Epub 2022 Feb 6. Thorac Cancer. 2022. PMID: 35128818 Free PMC article.
References
-
- Gottesman MM, Ambudkar SV, Ni B, Aran JM, Sugimoto Y, Cardarelli CO, Pastan I. Exploiting multidrug resistance to treat cancer. Cold Spring Harb Symp on Quant Biol. 1994;59:667–683. - PubMed
-
- Hannon MJ. Metal-based anticancer drugs: From a poast anchored in platinum chemistry to a post-genomic future of diverse chemistry and biology. Pure Appl Chem. 2007;79:2261.
-
- Minotti G, Menna P, Salvatorelli E, Cairo G, Gianni L. Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol Rev. 2004;56:185–229. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
