

ATPase site architecture is required for self-assembly and remodeling activity of a hexameric AAA+ transcriptional activator
- PMID: 22789710
- PMCID: PMC3419264
- DOI: 10.1016/j.molcel.2012.06.012
ATPase site architecture is required for self-assembly and remodeling activity of a hexameric AAA+ transcriptional activator
Abstract
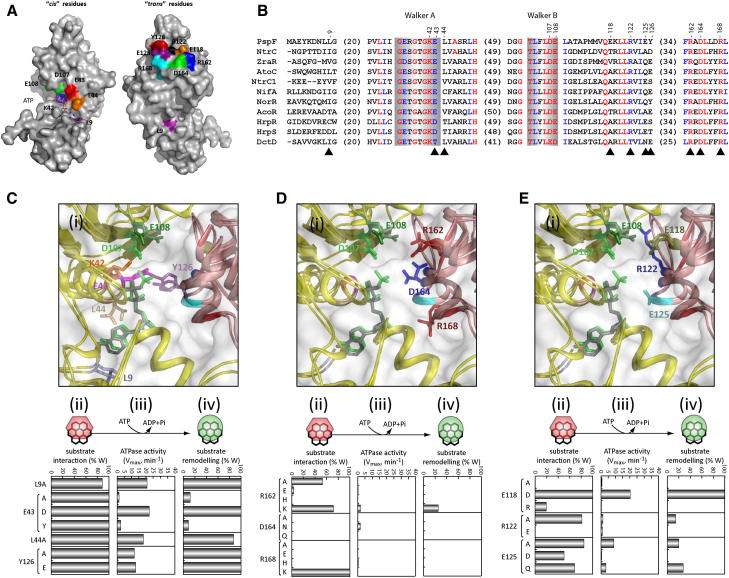
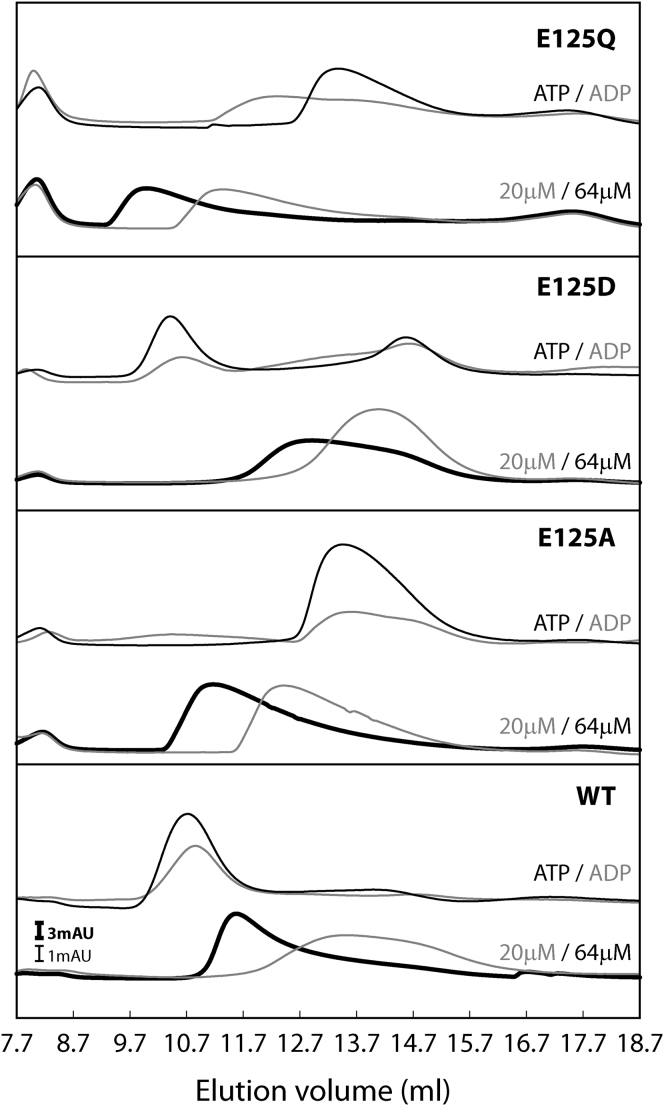
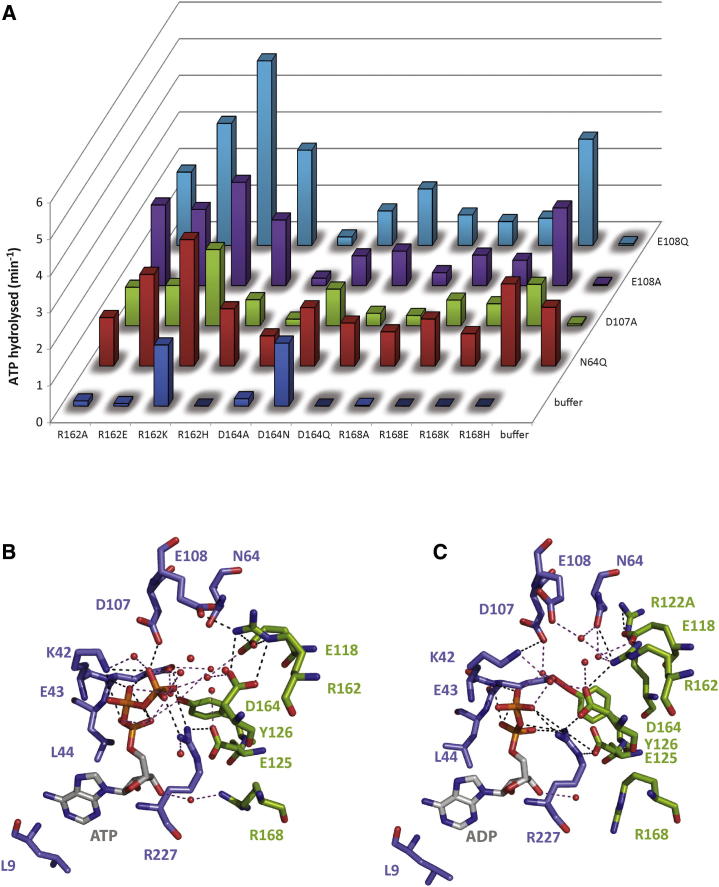
AAA+ proteins (ATPases associated with various cellular activities) are oligomeric ATPases that use ATP hydrolysis to remodel their substrates. By similarity with GTPases, a dynamic organization of the nucleotide-binding pockets between ATPase protomers is proposed to regulate functionality. Using the transcription activator PspF as an AAA+ model, we investigated contributions of conserved residues for roles in ATP hydrolysis and intersubunit communication. We determined the R-finger residue and revealed that it resides in a conserved "R-hand" motif (R(x)D(xxx)R) needed for its "trans-acting" activity. Further, a divergent Walker A glutamic acid residue acts synergistically with a tyrosine residue to function in ADP-dependent subunit-subunit coordination, forming the "ADP-switch" motif. Another glutamic acid controls hexamer formation in the presence of nucleotides. Together, these results lead to a "residue-nucleotide" interaction map upon which to base AAA+ core regulation.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Sigma54-dependent transcription activator phage shock protein F of Escherichia coli: a fragmentation approach to identify sequences that contribute to self-association.Biochem J. 2004 Mar 15;378(Pt 3):735-44. doi: 10.1042/BJ20031464. Biochem J. 2004. PMID: 14659000 Free PMC article.
-
Mechanism of homotropic control to coordinate hydrolysis in a hexameric AAA+ ring ATPase.J Mol Biol. 2008 Aug 1;381(1):1-12. doi: 10.1016/j.jmb.2008.05.075. Epub 2008 Jun 5. J Mol Biol. 2008. PMID: 18599077
-
An intramolecular route for coupling ATPase activity in AAA+ proteins for transcription activation.J Biol Chem. 2008 May 16;283(20):13725-35. doi: 10.1074/jbc.M800801200. Epub 2008 Mar 6. J Biol Chem. 2008. PMID: 18326037
-
Coupling AAA protein function to regulated gene expression.Biochim Biophys Acta. 2012 Jan;1823(1):108-16. doi: 10.1016/j.bbamcr.2011.08.012. Epub 2011 Aug 31. Biochim Biophys Acta. 2012. PMID: 21906631 Review.
-
MinD and role of the deviant Walker A motif, dimerization and membrane binding in oscillation.Mol Microbiol. 2003 Apr;48(2):295-303. doi: 10.1046/j.1365-2958.2003.03427.x. Mol Microbiol. 2003. PMID: 12675792 Review.
Cited by
-
An Arginine Finger Regulates the Sequential Action of Asymmetrical Hexameric ATPase in the Double-Stranded DNA Translocation Motor.Mol Cell Biol. 2016 Sep 12;36(19):2514-23. doi: 10.1128/MCB.00142-16. Print 2016 Oct 1. Mol Cell Biol. 2016. PMID: 27457616 Free PMC article.
-
A key hydrophobic patch identified in an AAA⁺ protein essential for its in trans inhibitory regulation.J Mol Biol. 2013 Aug 9;425(15):2656-69. doi: 10.1016/j.jmb.2013.04.024. Epub 2013 May 7. J Mol Biol. 2013. PMID: 23659791 Free PMC article.
-
The bacterial enhancer-dependent RNA polymerase.Biochem J. 2016 Nov 1;473(21):3741-3753. doi: 10.1042/BCJ20160741C. Biochem J. 2016. PMID: 27789741 Free PMC article. Review.
-
Engineered CRISPRa enables programmable eukaryote-like gene activation in bacteria.Nat Commun. 2019 Aug 26;10(1):3693. doi: 10.1038/s41467-019-11479-0. Nat Commun. 2019. PMID: 31451697 Free PMC article.
-
Assessing heterogeneity in oligomeric AAA+ machines.Cell Mol Life Sci. 2017 Mar;74(6):1001-1018. doi: 10.1007/s00018-016-2374-z. Epub 2016 Sep 26. Cell Mol Life Sci. 2017. PMID: 27669691 Free PMC article. Review.
References
-
- Ahmadian M.R., Stege P., Scheffzek K., Wittinghofer A. Confirmation of the arginine-finger hypothesis for the GAP-stimulated GTP-hydrolysis reaction of Ras. Nat. Struct. Biol. 1997;4:686–689. - PubMed
-
- Besche H., Tamura N., Tamura T., Zwickl P. Mutational analysis of conserved AAA+ residues in the archaeal Lon protease from Thermoplasma acidophilum. FEBS Lett. 2004;574:161–166. - PubMed
-
- Bochtler M., Hartmann C., Song H.K., Bourenkov G.P., Bartunik H.D., Huber R. The structures of HsIU and the ATP-dependent protease HsIU-HsIV. Nature. 2000;403:800–805. - PubMed
-
- Cannon W.V., Gallegos M.T., Buck M. Isomerization of a binary sigma-promoter DNA complex by transcription activators. Nat. Struct. Biol. 2000;7:594–601. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
