

Mechanical properties of lipid bilayers and regulation of mechanosensitive function: from biological to biomimetic channels
- PMID: 22790280
- PMCID: PMC3508901
- DOI: 10.4161/chan.21085
Mechanical properties of lipid bilayers and regulation of mechanosensitive function: from biological to biomimetic channels
Abstract
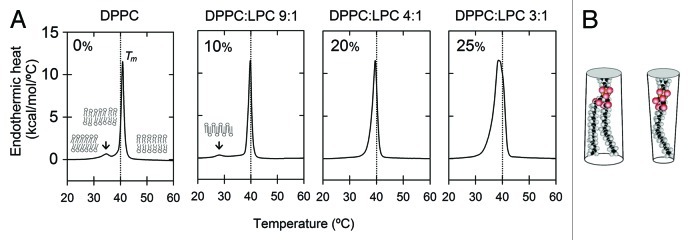
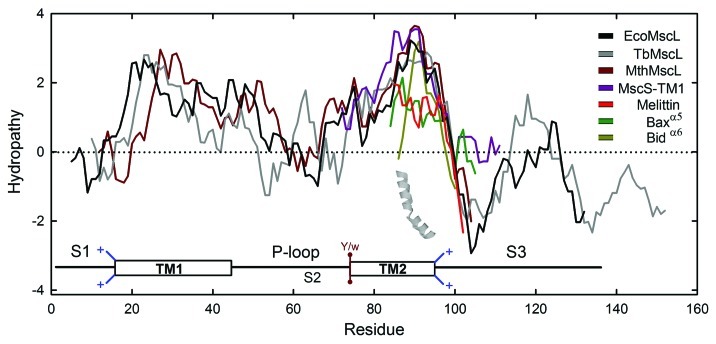
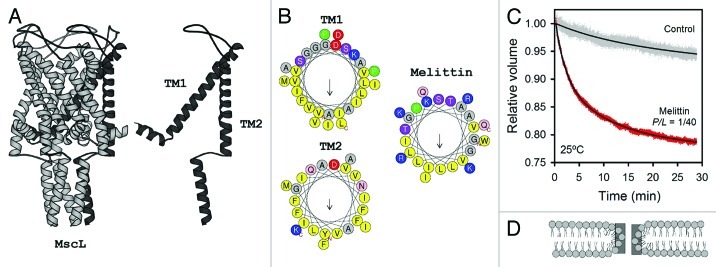
Material properties of lipid bilayers, including thickness, intrinsic curvature and compressibility regulate the function of mechanosensitive (MS) channels. This regulation is dependent on phospholipid composition, lateral packing and organization within the membrane. Therefore, a more complete framework to understand the functioning of MS channels requires insights into bilayer structure, thermodynamics and phospholipid structure, as well as lipid-protein interactions. Phospholipids and MS channels interact with each other mainly through electrostatic forces and hydrophobic matching, which are also crucial for antimicrobial peptides. They are excellent models for studying the formation and stabilization of membrane pores. Importantly, they perform equivalent responses as MS channels: (1) tilting in response to tension and (2) dissipation of osmotic gradients. Lessons learned from pore forming peptides could enrich our knowledge of mechanisms of action and evolution of these channels. Here, the current state of the art is presented and general principles of membrane regulation of mechanosensitive function are discussed.
Figures

References
-
- Martinac B. 3.5 Billion years of mechanosensory transduction: structure and function of mechanosensitive channels in prokaryotes. Current Topics Membr. 2007;58:25–57. doi: 10.1016/S1063-5823(06)58002-0. - DOI
-
- Levina N, Tötemeyer S, Stokes NR, Louis P, Jones MA, Booth IR. Protection of Escherichia coli cells against extreme turgor by activation of MscS and MscL mechanosensitive channels: identification of genes required for MscS activity. EMBO J. 1999;18:1730–7. doi: 10.1093/emboj/18.7.1730. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources