

Gating the mechanical channel Piezo1: a comparison between whole-cell and patch recording
- PMID: 22790451
- PMCID: PMC3508907
- DOI: 10.4161/chan.21064
Gating the mechanical channel Piezo1: a comparison between whole-cell and patch recording
Abstract
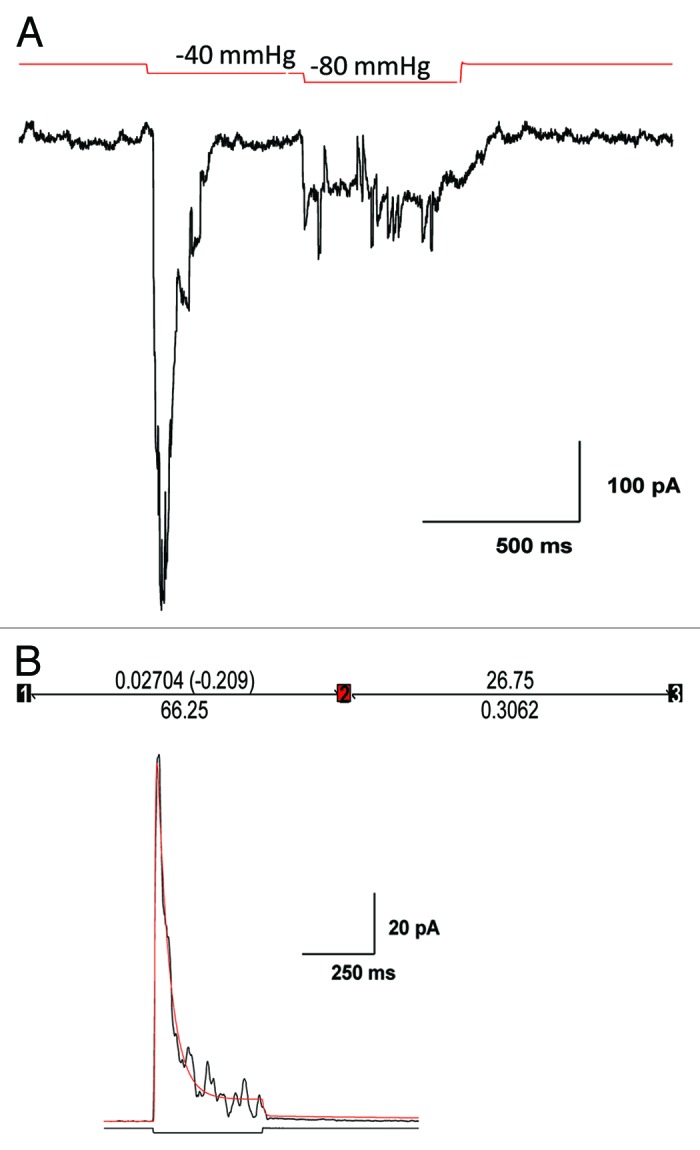
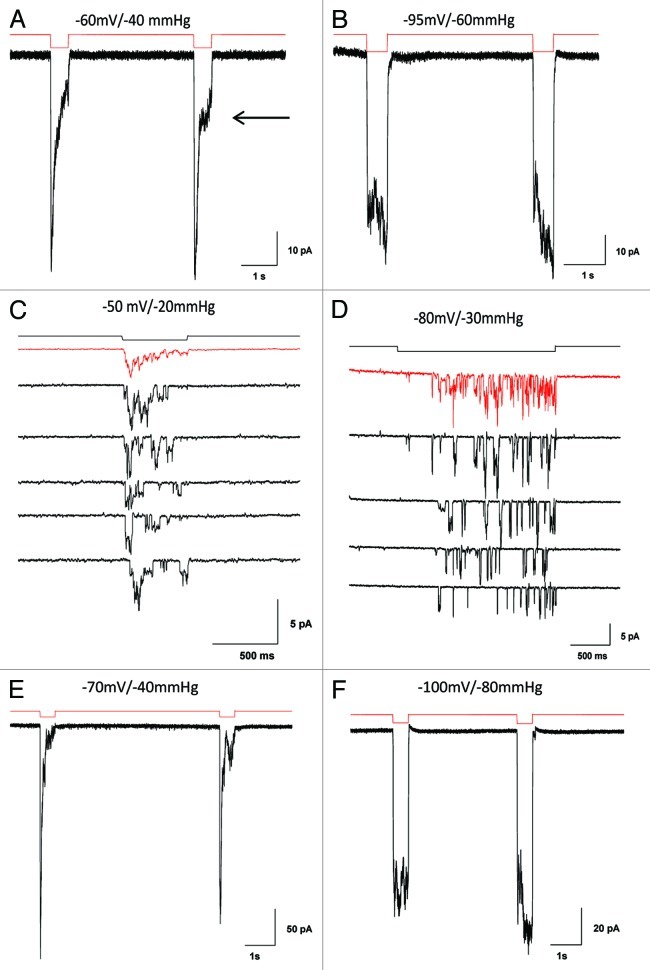
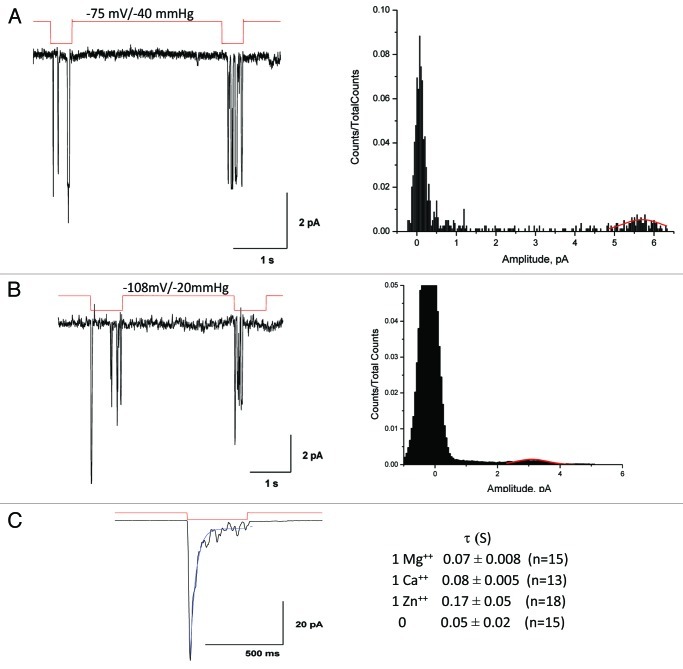
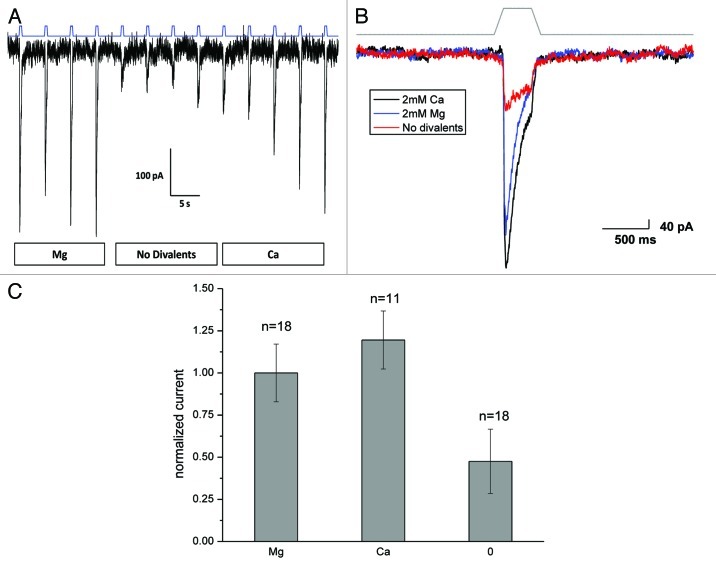
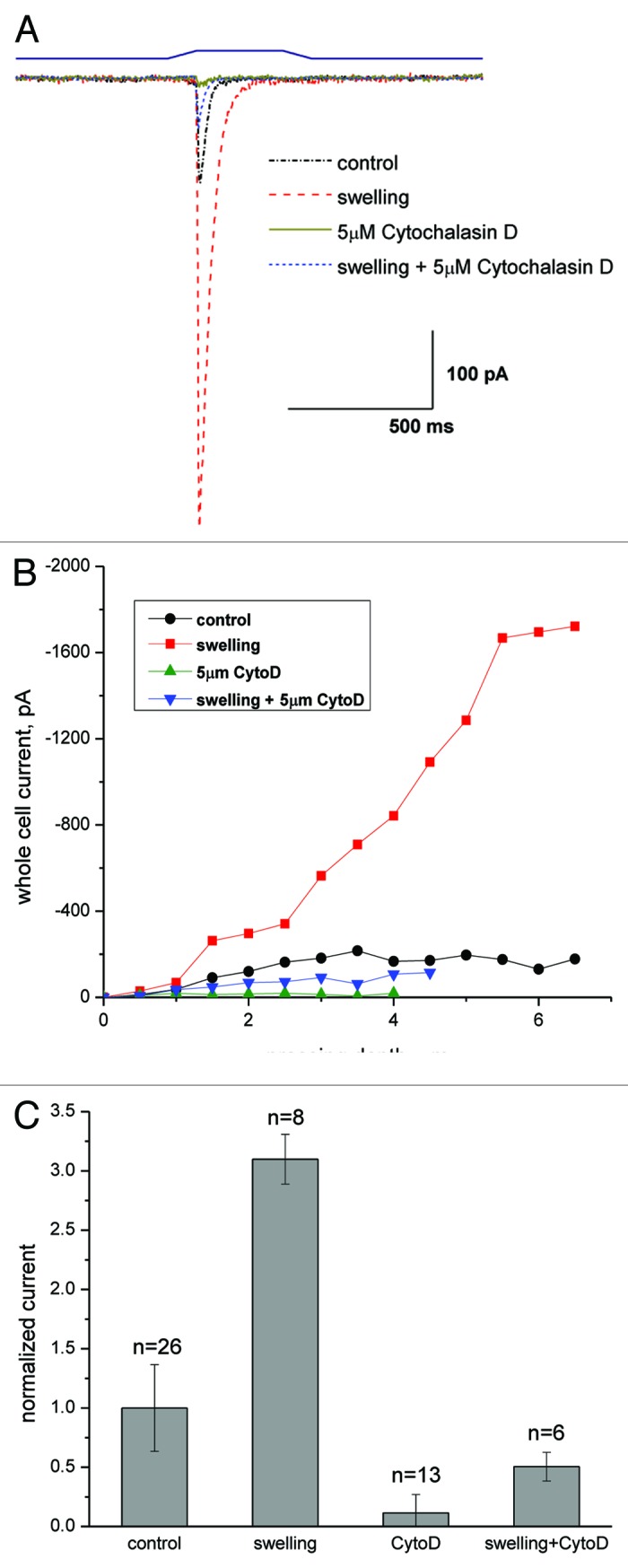
Piezo1 is a eukaryotic cation-selective mechanosensitive ion channel. To understand channel function in vivo, we first need to analyze and compare the response in the whole cell and the patch. In patches, Piezo1 inactivates and the current is fit well by a 3-state model with a single pressure-dependent rate. However, repeated stimulation led to an irreversible loss of inactivation. Remarkably, the loss of inactivation did not occur on a channel-by-channel basis but on all channels at the same time. Thus, the channels are in common mechanical domain. Divalent ions decreased the unitary conductance from ~68 pS to ~37 pS, irrespective of the cation species. Mg and Ca did not affect inactivation rates, but Zn caused a 3-fold slowing. CytochalasinD (cytoD) does not alter inactivation rates or the transition to the non-inactivating mode but does reduce the steady-state response. Whole-cell currents were similar to patch currents but also had significant differences. In contrast to the patch, cytoD inhibited the current suggesting that the activating forces were transmitted through the actin cytoskeleton. Hypotonic swelling that prestressed the cytoskeleton and the bilayer greatly increased the sensitivity of both control and cytoD cells so there are two pathways to transmit force to the channels. In contrast to patch, removing divalent ions decreased the whole-cell current. The difference between whole cell and patch properties provide new insights into our understanding of the Piezo1 gating mechanisms and cautions against generalization to in situ behavior.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources