

Interaction of temperature and irradiance effects on photosynthetic acclimation in two accessions of Arabidopsis thaliana
- PMID: 22791015
- PMCID: PMC3430840
- DOI: 10.1007/s11120-012-9756-3
Interaction of temperature and irradiance effects on photosynthetic acclimation in two accessions of Arabidopsis thaliana
Abstract
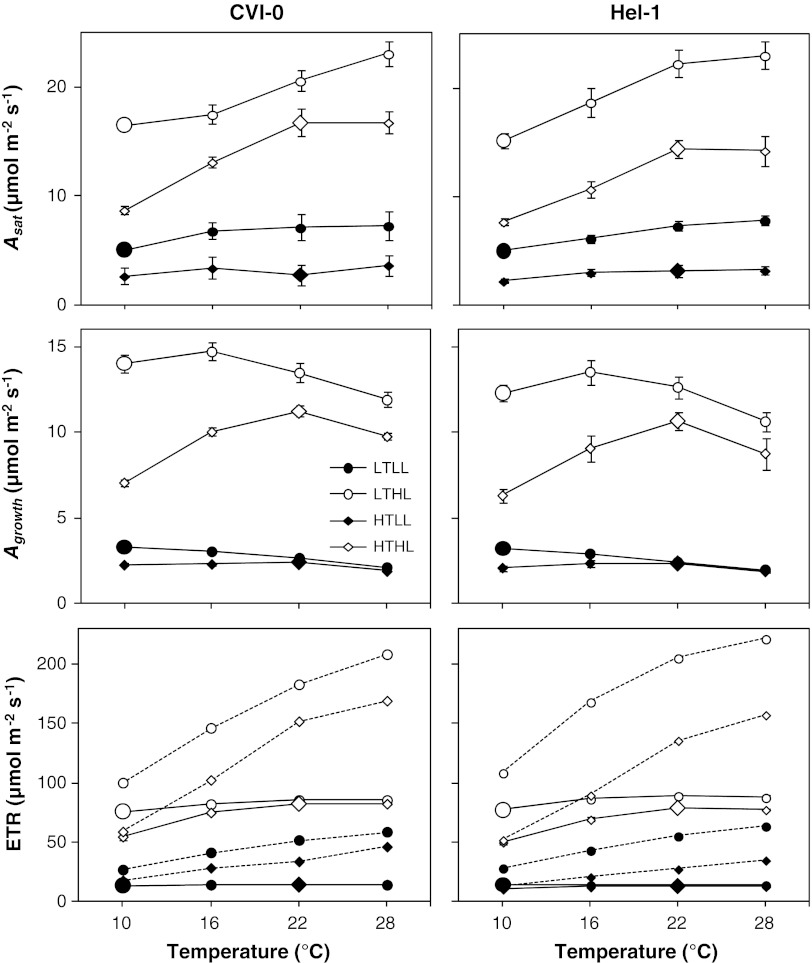
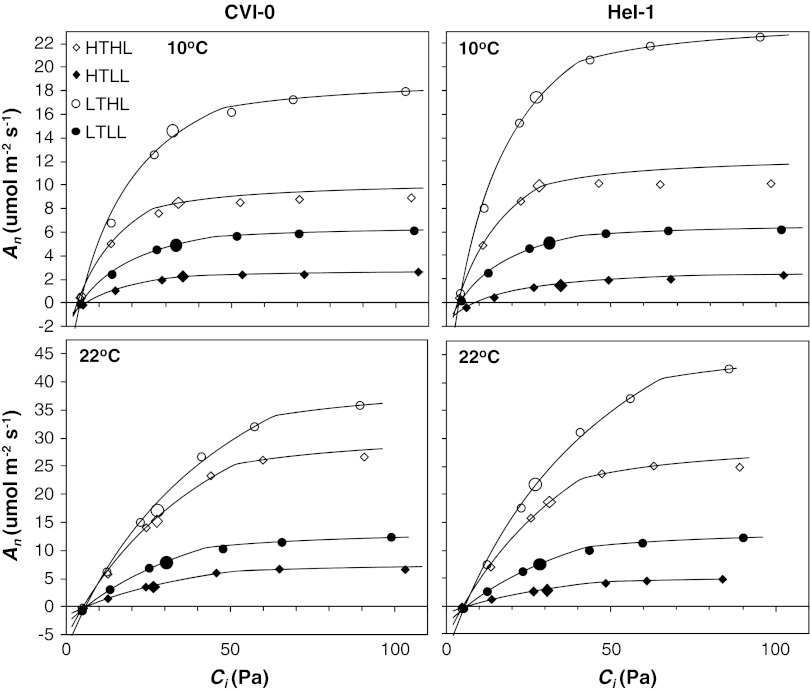
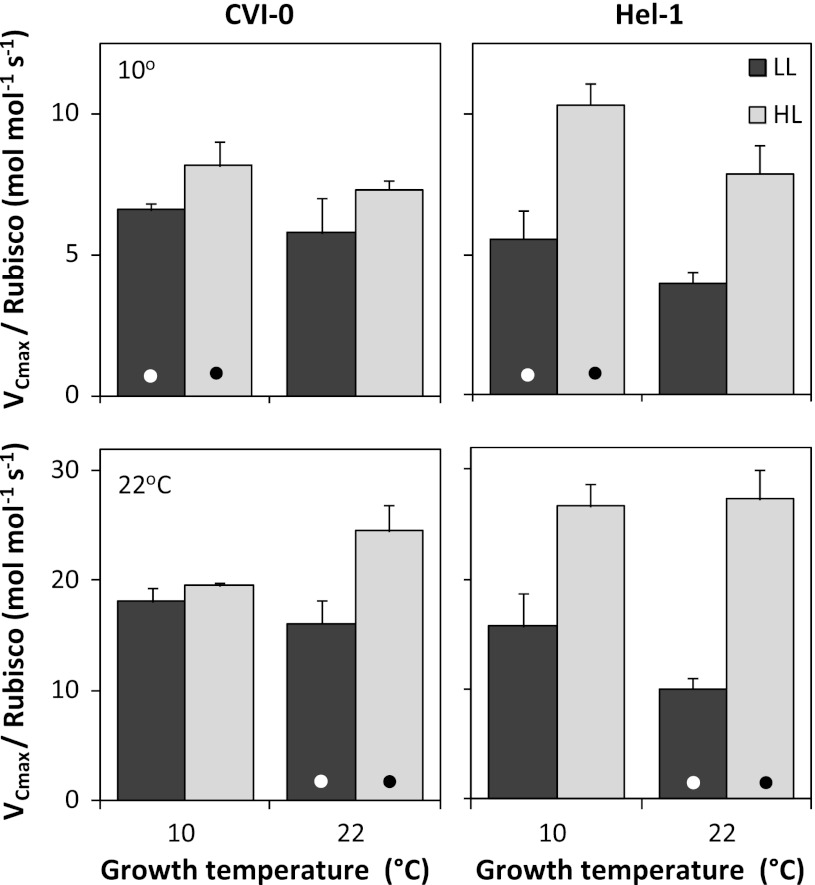
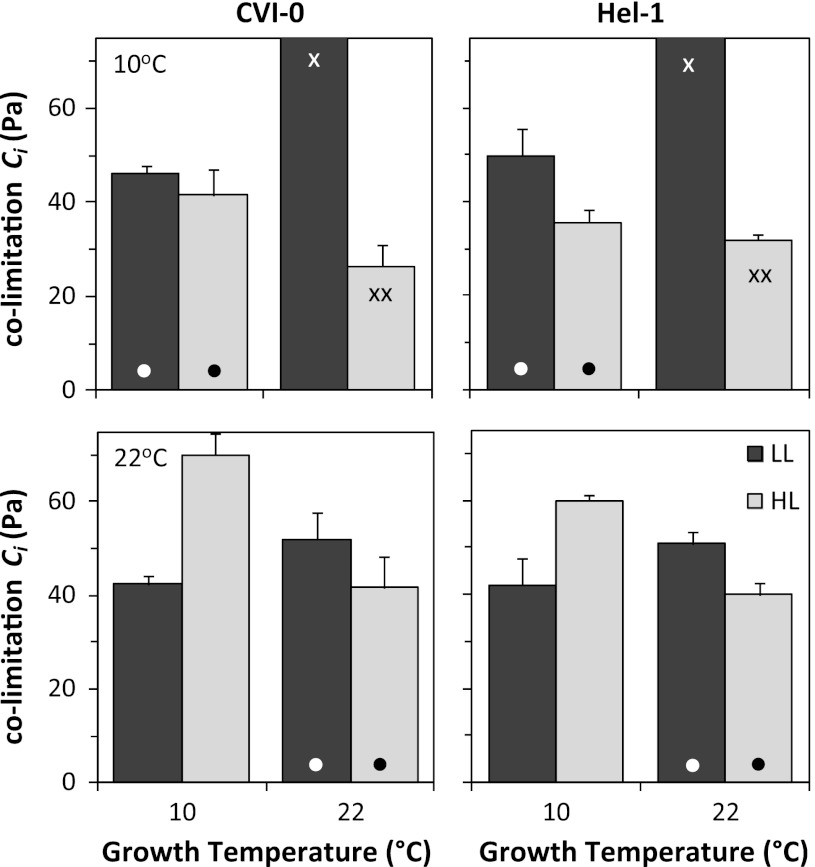
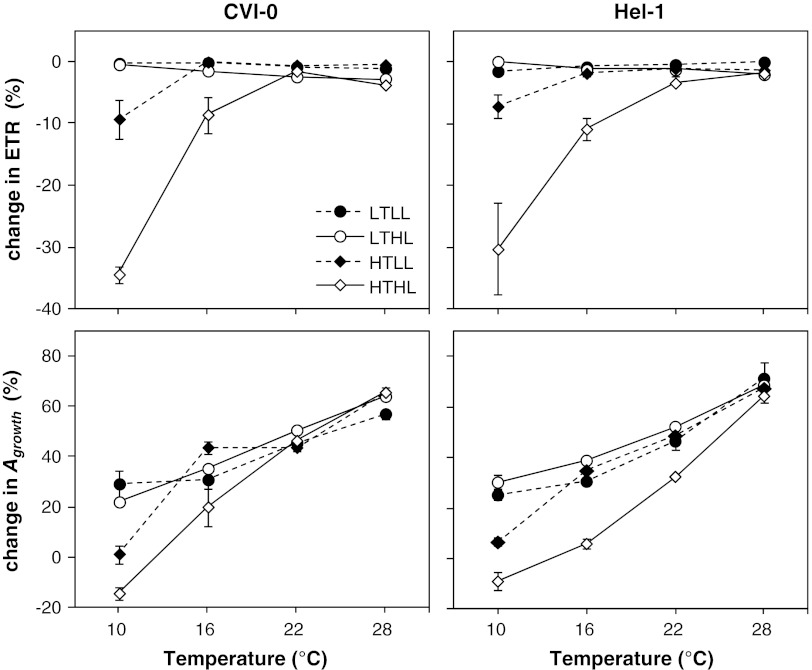
The effect of temperature and irradiance during growth on photosynthetic traits of two accessions of Arabidopsis thaliana was investigated. Plants were grown at 10 and 22 °C, and at 50 and 300 μmol photons m(-2) s(-1) in a factorial design. As known from other cold-tolerant herbaceous species, growth of Arabidopsis at low temperature resulted in increases in photosynthetic capacity per unit leaf area and chlorophyll. Growth at high irradiance had a similar effect. However, the growth temperature and irradiance showed interacting effects for several capacity-related variables. Temperature effects on the ratio between electron transport capacity and carboxylation capacity were also different in low compared to high irradiance grown Arabidopsis. The carboxylation capacity per unit Rubisco, a measure for the in vivo Rubisco activity, was low in low irradiance grown plants but there was no clear growth temperature effect. The limitation of photosynthesis by the utilization of triose-phosphate in high temperature grown plants was less when grown at low compared to high irradiance. Several of these traits contribute to reduced efficiency of the utilization of resources for photosynthesis of Arabidopsis at low irradiance. The two accessions from contrasting climates showed remarkably similar capabilities of developmental acclimation to the two environmental factors. Hence, no evidence was found for photosynthetic adaptation of the photosynthetic apparatus to specific climatic conditions.
Figures
References
-
- Atkin OK, Scheurwater I, Pons TL. High thermal acclimation potential of both photosynthesis and respiration in two lowland Plantago species in contrast to an alpine congeneric. Global Change Biol. 2006;12:500–515. doi: 10.1111/j.1365-2486.2006.01114.x. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
