

Biochemical characterization of a recombinant SARS coronavirus nsp12 RNA-dependent RNA polymerase capable of copying viral RNA templates
- PMID: 22791111
- PMCID: PMC7086750
- DOI: 10.1007/s00705-012-1404-x
Biochemical characterization of a recombinant SARS coronavirus nsp12 RNA-dependent RNA polymerase capable of copying viral RNA templates
Abstract
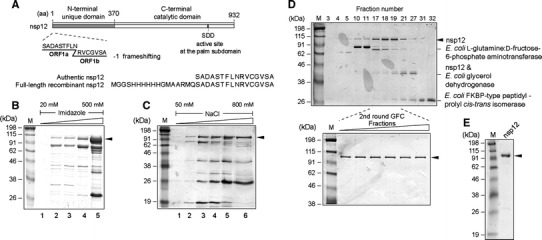
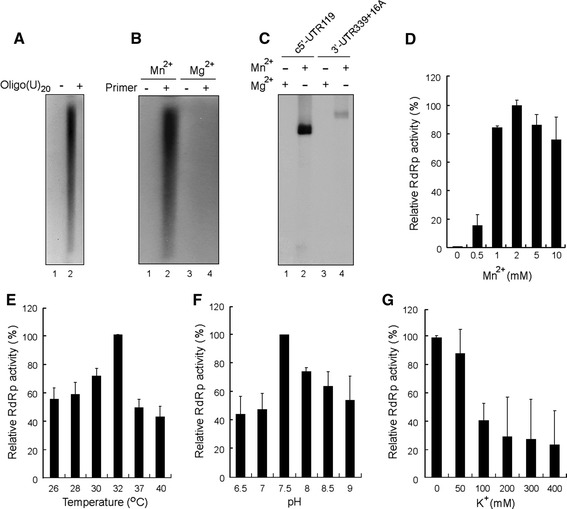
The severe acute respiratory syndrome coronavirus (SARS-CoV) RNA genome is replicated by a virus-encoded RNA replicase, the key component of which is the nonstructural protein 12 (nsp12). In this report, we describe the biochemical properties of a full-length recombinant SARS-CoV nsp12 RNA-dependent RNA polymerase (RdRp) capable of copying viral RNA templates. The purified SARS-CoV nsp12 showed both primer-dependent and primer-independent RNA synthesis activities using homopolymeric RNA templates. The RdRp activity was strictly dependent on Mn(2+). The nsp12 preferentially copied homopolymeric pyrimidine RNA templates in the absence of an added oligonucleotide primer. It was also able to initiate de novo RNA synthesis from the 3'-ends of both the plus- and minus-strand genome of SARS-CoV, using the 3'-terminal 36- and 37-nt RNA, respectively. The in vitro RdRp assay system established with a full-length nsp12 will be useful for understanding the mechanisms of coronavirus replication and for the development of anti-SARS-CoV agents.
Figures

References
-
- Clemente-Casares P, Lopez-Jimenez AJ, Bellon-Echeverria I, Encinar JA, Martinez-Alfaro E, Perez-Flores R, Mas A. De novo polymerase activity and oligomerization of hepatitis C virus RNA-dependent RNA-polymerases from genotypes 1 to 5. PLoS One. 2011;6:e18515. doi: 10.1371/journal.pone.0018515. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
