

Characterization of SOC1's central role in flowering by the identification of its upstream and downstream regulators
- PMID: 22791302
- PMCID: PMC3440217
- DOI: 10.1104/pp.112.202614
Characterization of SOC1's central role in flowering by the identification of its upstream and downstream regulators
Erratum in
- Plant Physiol. 2013 Aug;162(4):2151
Abstract
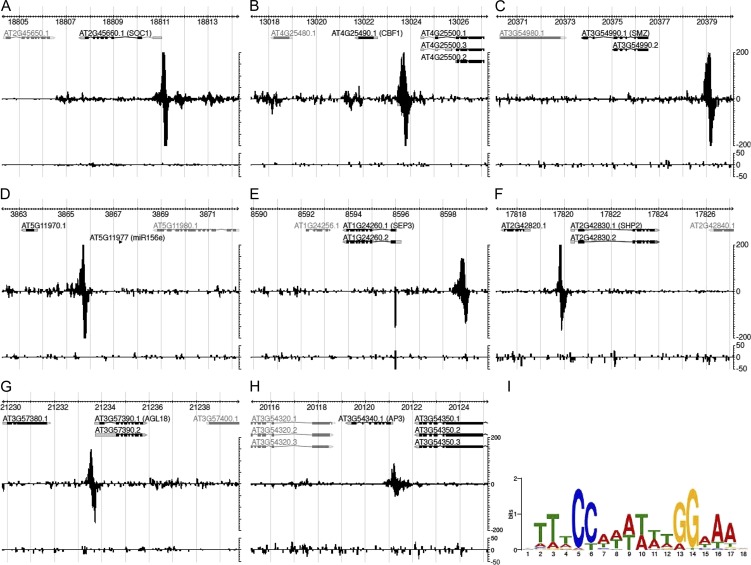
The transition from vegetative to reproductive development is one of the most important phase changes in the plant life cycle. This step is controlled by various environmental signals that are integrated at the molecular level by so-called floral integrators. One such floral integrator in Arabidopsis (Arabidopsis thaliana) is the MADS domain transcription factor SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (SOC1). Despite extensive genetic studies, little is known about the transcriptional control of SOC1, and we are just starting to explore the network of genes under the direct control of SOC1 transcription factor complexes. Here, we show that several MADS domain proteins, including SOC1 heterodimers, are able to bind SOC1 regulatory sequences. Genome-wide target gene analysis by ChIP-seq confirmed the binding of SOC1 to its own locus and shows that it also binds to a plethora of flowering-time regulatory and floral homeotic genes. In turn, the encoded floral homeotic MADS domain proteins appear to bind SOC1 regulatory sequences. Subsequent in planta analyses revealed SOC1 repression by several floral homeotic MADS domain proteins, and we show that, mechanistically, this depends on the presence of the SOC1 protein. Together, our data show that SOC1 constitutes a major hub in the regulatory networks underlying floral timing and flower development and that these networks are composed of many positive and negative autoregulatory and feedback loops. The latter seems to be crucial for the generation of a robust flower-inducing signal, followed shortly after by repression of the SOC1 floral integrator.
Figures





References
-
- Adamczyk BJ, Lehti-Shiu MD, Fernandez DE. (2007) The MADS domain factors AGL15 and AGL18 act redundantly as repressors of the floral transition in Arabidopsis. Plant J 50: 1007–1019 - PubMed
-
- Alon U. (2007) Network motifs: theory and experimental approaches. Nat Rev Genet 8: 450–461 - PubMed
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Benlloch R, Kim MC, Sayou C, Thevenon E, Parcy F, Nilsson O. (2011) Integrating long-day flowering signals: a LEAFY binding site is essential for proper photoperiodic activation of APETALA1. Plant J 67: 1094–1102 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
