

The sensitive to freezing3 mutation of Arabidopsis thaliana is a cold-sensitive allele of homomeric acetyl-CoA carboxylase that results in cold-induced cuticle deficiencies
- PMID: 22791831
- PMCID: PMC3431002
- DOI: 10.1093/jxb/ers191
The sensitive to freezing3 mutation of Arabidopsis thaliana is a cold-sensitive allele of homomeric acetyl-CoA carboxylase that results in cold-induced cuticle deficiencies
Abstract
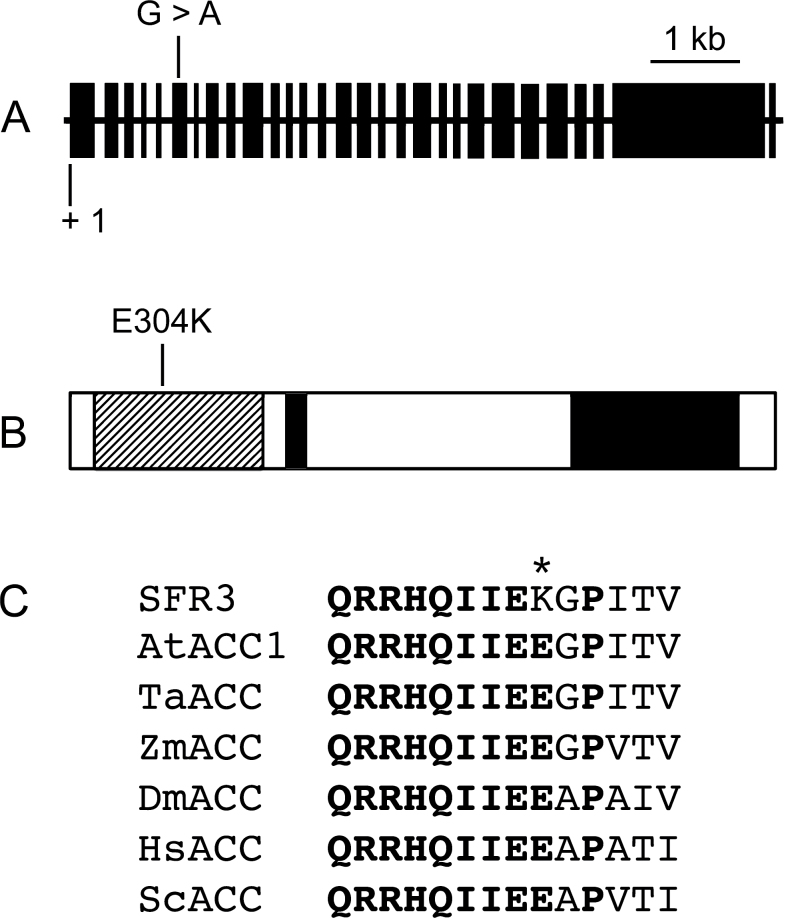
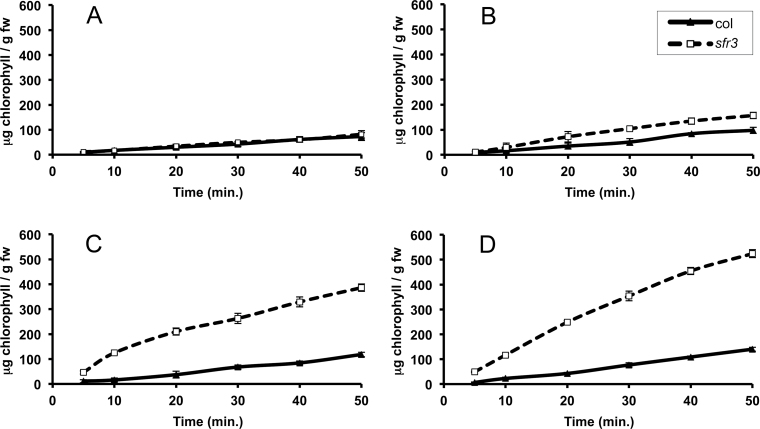
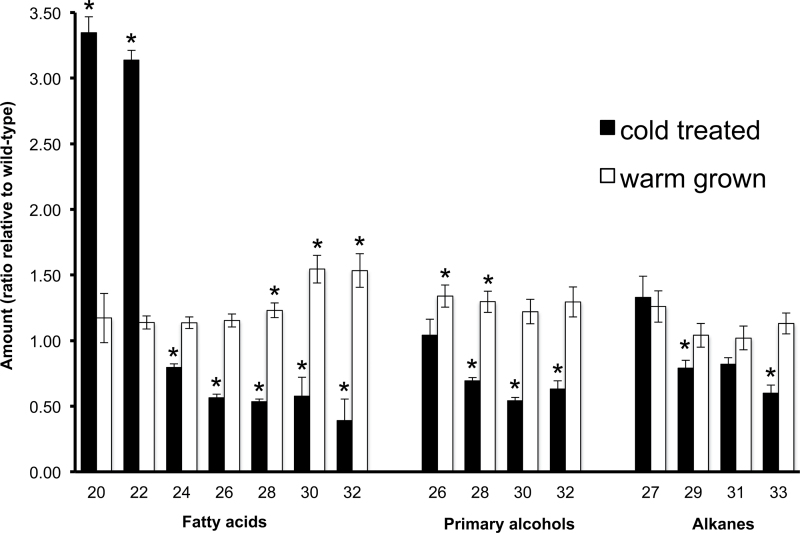
The sfr3 mutation causes freezing sensitivity in Arabidopsis thaliana. Mapping, sequencing, and transgenic complementation showed sfr3 to be a missense mutation in ACC1, an essential gene encoding homomeric (multifunctional) acetyl-CoA carboxylase. Cuticle permeability was compromised in the sfr3 mutant when plants were grown in the cold but not in the warm. Wax deposition on the inflorescence stem of cold-grown sfr3 plants was inhibited and the long-chain components of their leaf cuticular wax were reduced compared with wild-type plants. Thus, freezing sensitivity of sfr3 appears, from these results, to be due to cuticular deficiencies that develop during cold acclimation. These observations demonstrated the essential role of the cuticle in tolerance to freezing and drought.
Figures




Similar articles
-
The glossyhead1 allele of ACC1 reveals a principal role for multidomain acetyl-coenzyme A carboxylase in the biosynthesis of cuticular waxes by Arabidopsis.Plant Physiol. 2011 Nov;157(3):1079-92. doi: 10.1104/pp.111.185132. Epub 2011 Sep 23. Plant Physiol. 2011. PMID: 21949210 Free PMC article.
-
Dissecting the Roles of Cuticular Wax in Plant Resistance to Shoot Dehydration and Low-Temperature Stress in Arabidopsis.Int J Mol Sci. 2021 Feb 4;22(4):1554. doi: 10.3390/ijms22041554. Int J Mol Sci. 2021. PMID: 33557073 Free PMC article.
-
Identification of ICE2, a gene involved in cold acclimation which determines freezing tolerance in Arabidopsis thaliana.Gene. 2009 Jan 15;429(1-2):98-103. doi: 10.1016/j.gene.2008.10.016. Epub 2008 Nov 5. Gene. 2009. PMID: 19026725
-
Nitric reductase-dependent nitric oxide production is involved in cold acclimation and freezing tolerance in Arabidopsis.Plant Physiol. 2009 Oct;151(2):755-67. doi: 10.1104/pp.109.140996. Epub 2009 Aug 26. Plant Physiol. 2009. PMID: 19710235 Free PMC article.
-
Cold acclimation by the CBF-COR pathway in a changing climate: Lessons from Arabidopsis thaliana.Plant Cell Rep. 2019 May;38(5):511-519. doi: 10.1007/s00299-019-02376-3. Epub 2019 Jan 16. Plant Cell Rep. 2019. PMID: 30652229 Free PMC article. Review.
Cited by
-
The Acetate Pathway Supports Flavonoid and Lipid Biosynthesis in Arabidopsis.Plant Physiol. 2020 Feb;182(2):857-869. doi: 10.1104/pp.19.00683. Epub 2019 Nov 12. Plant Physiol. 2020. PMID: 31719153 Free PMC article.
-
Overexpression of a novel cold-responsive transcript factor LcFIN1 from sheepgrass enhances tolerance to low temperature stress in transgenic plants.Plant Biotechnol J. 2016 Mar;14(3):861-74. doi: 10.1111/pbi.12435. Epub 2015 Aug 3. Plant Biotechnol J. 2016. PMID: 26234381 Free PMC article.
-
MUR1-mediated cell-wall fucosylation is required for freezing tolerance in Arabidopsis thaliana.New Phytol. 2019 Dec;224(4):1518-1531. doi: 10.1111/nph.16209. New Phytol. 2019. PMID: 31549420 Free PMC article.
-
Constitutive and Chloroplast Targeted Expression of Acetyl-CoA Carboxylase in Oleaginous Microalgae Elevates Fatty Acid Biosynthesis.Mar Biotechnol (NY). 2018 Oct;20(5):566-572. doi: 10.1007/s10126-018-9841-5. Epub 2018 Jun 22. Mar Biotechnol (NY). 2018. PMID: 29931608
-
Structure, Assembly and Function of Cuticle from Mechanical Perspective with Special Focus on Perianth.Int J Mol Sci. 2021 Apr 16;22(8):4160. doi: 10.3390/ijms22084160. Int J Mol Sci. 2021. PMID: 33923850 Free PMC article. Review.
References
-
- Baud S, Guyon V, Kronenberger J, Wuilleme S, Miquel M, Caboche M, Lepiniec L, Rochat C. Multifunctional acetyl-CoA carboxylase 1 is essential for very long chain fatty acid elongation and embryo development in Arabidopsis. The Plant Journal. 2003;33:75–86. - PubMed
-
- Bell CJ, Ecker JR. Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics. 1994;19:137–144. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
