

Bringing together evolution on serpentine and polyploidy: spatiotemporal history of the diploid-tetraploid complex of Knautia arvensis (Dipsacaceae)
- PMID: 22792207
- PMCID: PMC3390331
- DOI: 10.1371/journal.pone.0039988
Bringing together evolution on serpentine and polyploidy: spatiotemporal history of the diploid-tetraploid complex of Knautia arvensis (Dipsacaceae)
Abstract
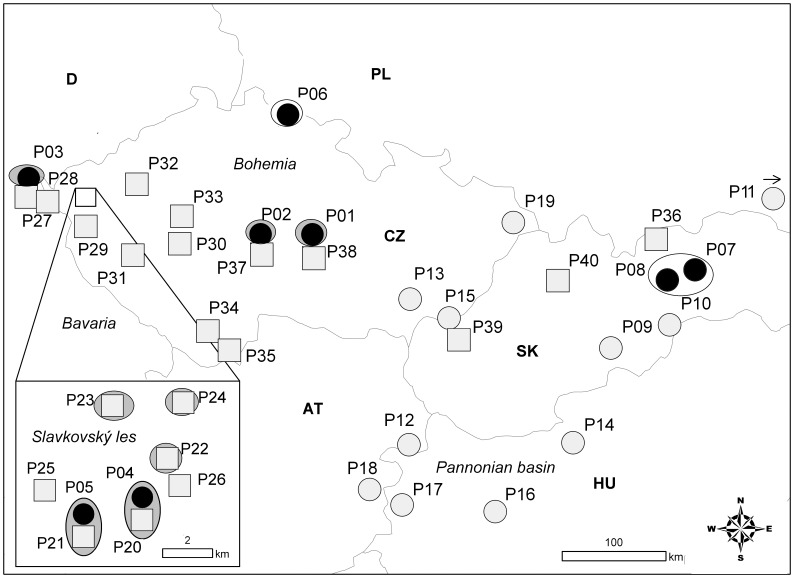
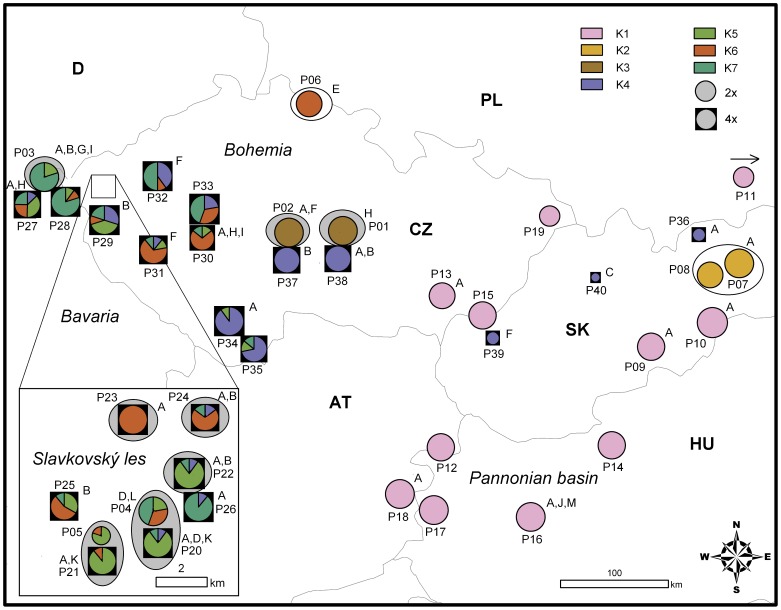
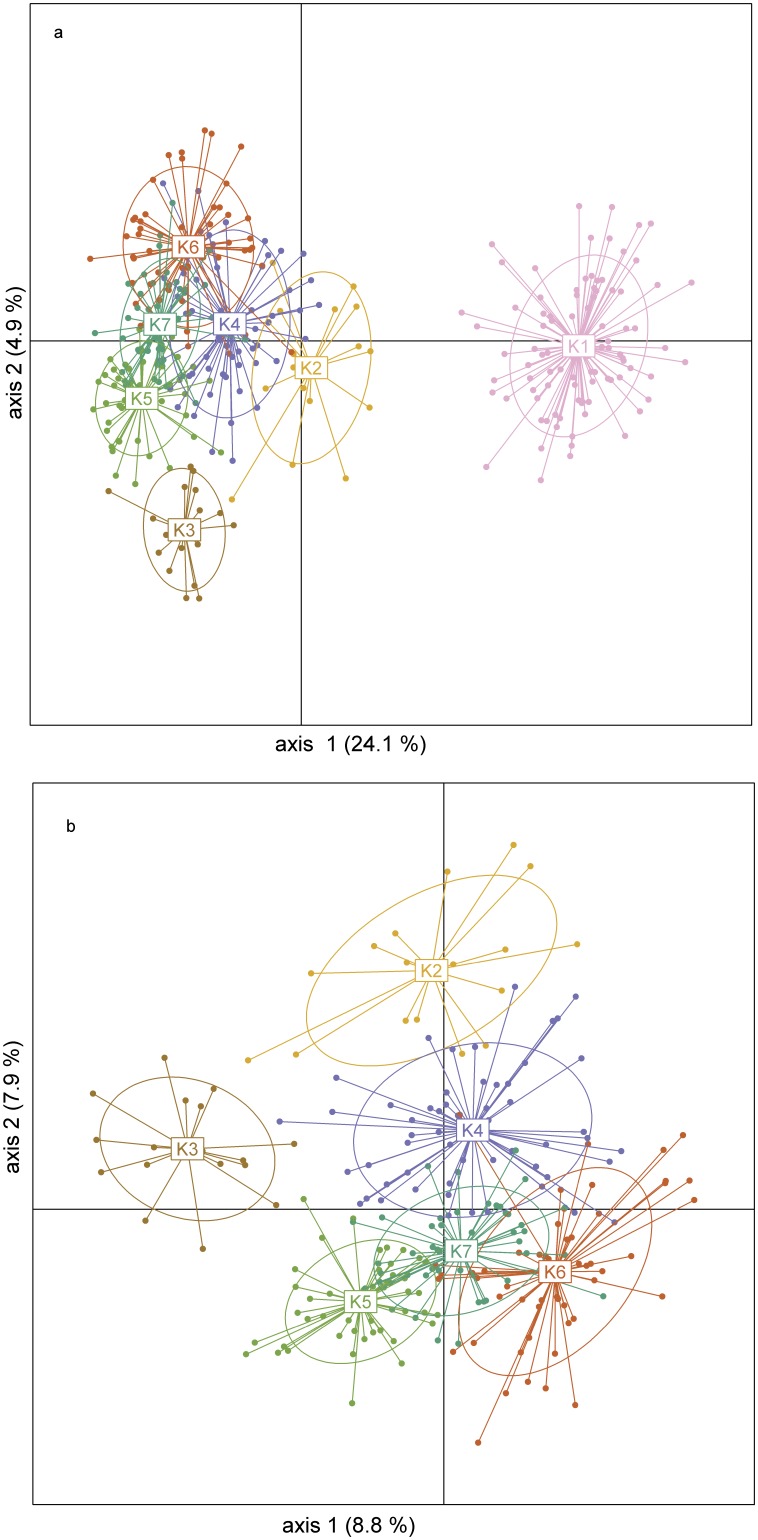
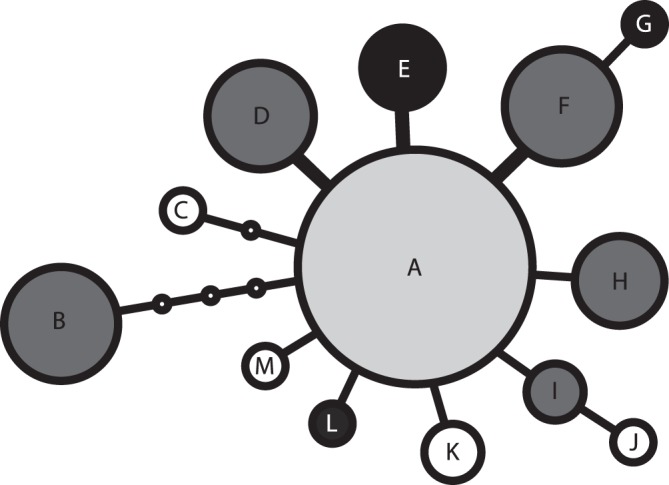
Polyploidization is one of the leading forces in the evolution of land plants, providing opportunities for instant speciation and rapid gain of evolutionary novelties. Highly selective conditions of serpentine environments act as an important evolutionary trigger that can be involved in various speciation processes. Whereas the significance of both edaphic speciation on serpentine and polyploidy is widely acknowledged in plant evolution, the links between polyploid evolution and serpentine differentiation have not yet been examined. To fill this gap, we investigated the evolutionary history of the perennial herb Knautia arvensis (Dipsacaceae), a diploid-tetraploid complex that exhibits an intriguing pattern of eco-geographic differentiation. Using plastid DNA sequencing and AFLP genotyping of 336 previously cytotyped individuals from 40 populations from central Europe, we unravelled the patterns of genetic variation among the cytotypes and the edaphic types. Diploids showed the highest levels of genetic differentiation, likely as a result of long term persistence of several lineages in ecologically distinct refugia and/or independent immigration. Recurrent polyploidization, recorded in one serpentine island, seems to have opened new possibilities for the local serpentine genotype. Unlike diploids, the serpentine tetraploids were able to escape from the serpentine refugium and spread further; this was also attributable to hybridization with the neighbouring non-serpentine tetraploid lineages. The spatiotemporal history of K. arvensis allows tracing the interplay of polyploid evolution and ecological divergence on serpentine, resulting in a complex evolutionary pattern. Isolated serpentine outcrops can act as evolutionary capacitors, preserving distinct karyological and genetic diversity. The serpentine lineages, however, may not represent evolutionary 'dead-ends' but rather dynamic systems with a potential to further influence the surrounding populations, e.g., via independent polyplodization and hybridization. The complex eco-geographical pattern together with the incidence of both primary and secondary diploid-tetraploid contact zones makes K. arvensis a unique system for addressing general questions of polyploid research.
Conflict of interest statement
Figures

Similar articles
-
Patterns of rapid diversification in heteroploid Knautia sect. Trichera (Caprifoliaceae, Dipsacoideae), one of the most intricate taxa of the European flora.BMC Evol Biol. 2016 Oct 10;16(1):204. doi: 10.1186/s12862-016-0773-2. BMC Evol Biol. 2016. PMID: 27724874 Free PMC article.
-
Disentangling relationships among the diploid members of the intricate genus Knautia (Caprifoliaceae, Dipsacoideae).Mol Phylogenet Evol. 2014 May;74:97-110. doi: 10.1016/j.ympev.2014.01.028. Epub 2014 Feb 6. Mol Phylogenet Evol. 2014. PMID: 24508604
-
Nonadaptive processes governing early stages of polyploid evolution: Insights from a primary contact zone of relict serpentine Knautia arvensis (Caprifoliaceae).Am J Bot. 2014 Jun 1;101(6):935-945. doi: 10.3732/ajb.1400005. Epub 2014 Jun 11. Am J Bot. 2014. PMID: 24920762
-
Polyploidy and interspecific hybridization: partners for adaptation, speciation and evolution in plants.Ann Bot. 2017 Aug 1;120(2):183-194. doi: 10.1093/aob/mcx079. Ann Bot. 2017. PMID: 28854567 Free PMC article. Review.
-
Factors promoting polyploid persistence and diversification and limiting diploid speciation during the K-Pg interlude.Curr Opin Plant Biol. 2018 Apr;42:1-7. doi: 10.1016/j.pbi.2017.09.010. Epub 2017 Oct 27. Curr Opin Plant Biol. 2018. PMID: 29107221 Review.
Cited by
-
De novo genome assembly and genome skims reveal LTRs dominate the genome of a limestone endemic Mountainsnail (Oreohelix idahoensis).BMC Genomics. 2022 Dec 2;23(1):796. doi: 10.1186/s12864-022-09000-x. BMC Genomics. 2022. PMID: 36460988 Free PMC article.
-
Epigenetic rather than genetic factors may explain phenotypic divergence between coastal populations of diploid and tetraploid Limonium spp. (Plumbaginaceae) in Portugal.BMC Plant Biol. 2013 Dec 6;13:205. doi: 10.1186/1471-2229-13-205. BMC Plant Biol. 2013. PMID: 24314092 Free PMC article.
-
The Evolutionary Genomics of Serpentine Adaptation.Front Plant Sci. 2020 Dec 16;11:574616. doi: 10.3389/fpls.2020.574616. eCollection 2020. Front Plant Sci. 2020. PMID: 33391295 Free PMC article. Review.
-
Evolutionary consequences, constraints and potential of polyploidy in plants.Cytogenet Genome Res. 2013;140(2-4):137-50. doi: 10.1159/000351727. Epub 2013 Jun 18. Cytogenet Genome Res. 2013. PMID: 23796571 Free PMC article. Review.
-
Evolution of nickel hyperaccumulation and serpentine adaptation in the Alyssum serpyllifolium species complex.Heredity (Edinb). 2017 Jan;118(1):31-41. doi: 10.1038/hdy.2016.93. Epub 2016 Oct 26. Heredity (Edinb). 2017. PMID: 27782119 Free PMC article.
References
-
- Brady KU, Kruckeberg AR, Bradshaw HD., Jr Evolutionary ecology of plant adaptation to serpentine soils. Annu Rev Ecol Evol Syst. 2005;36:243–266.
-
- Kazakou E, Dimitrakopoulos PG, Baker AJ, Reeves RD, Troumbis AY. Hypotheses, mechanisms and trade–offs of tolerance and adaptation to serpentine soils: from species to ecosystem level. Biol Rev Camb Philos Soc. 2008;83:495–508. - PubMed
-
- Proctor J. Toxins, nutrient shortages and droughts: the serpentine challenge. Trends Ecol Evol. 1999;14:334–335.
-
- Kruckeberg AR. California Serpentines: Flora, Vegetation, Geology, Soils and Management Problems. Berkeley: University of California Press. 1984.
-
- Kruckeberg AR. An essay: The stimulus of unusual geologies for plant speciation. Syst Bot. 1986;11:455–463.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
