

Estrogen leads to reversible hair cycle retardation through inducing premature catagen and maintaining telogen
- PMID: 22792225
- PMCID: PMC3390338
- DOI: 10.1371/journal.pone.0040124
Estrogen leads to reversible hair cycle retardation through inducing premature catagen and maintaining telogen
Abstract
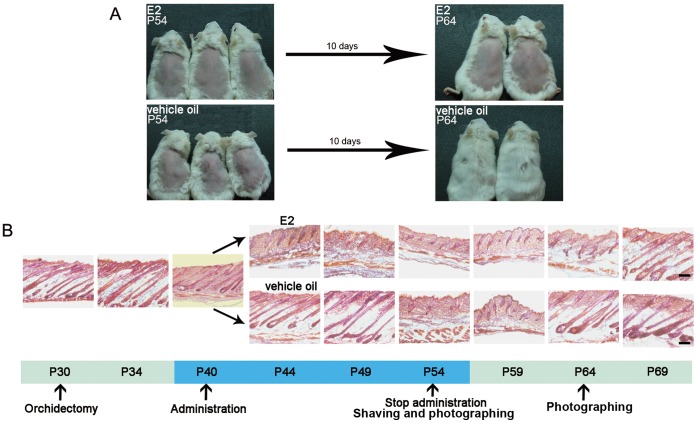
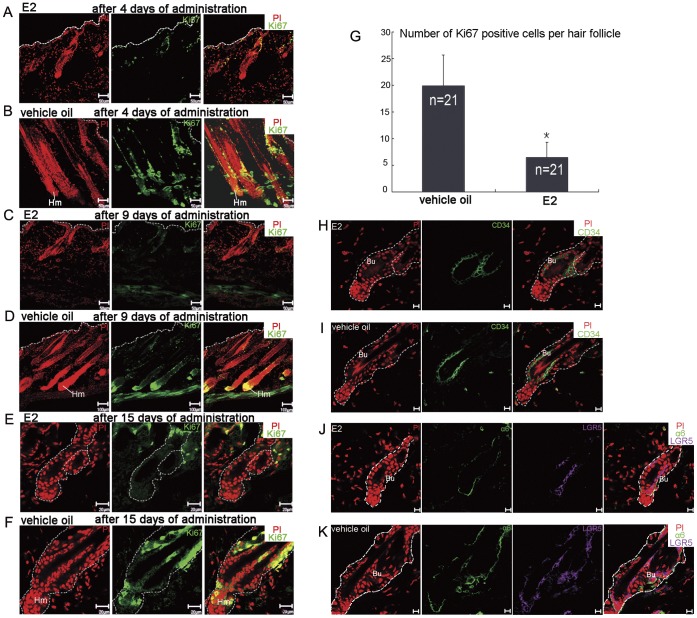
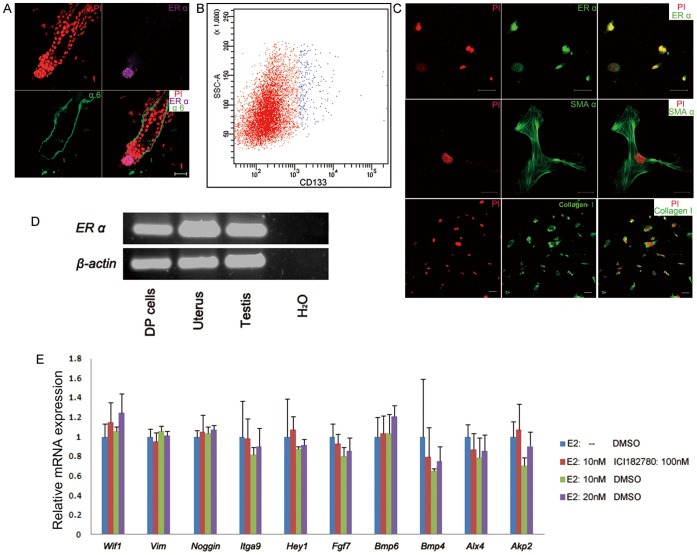
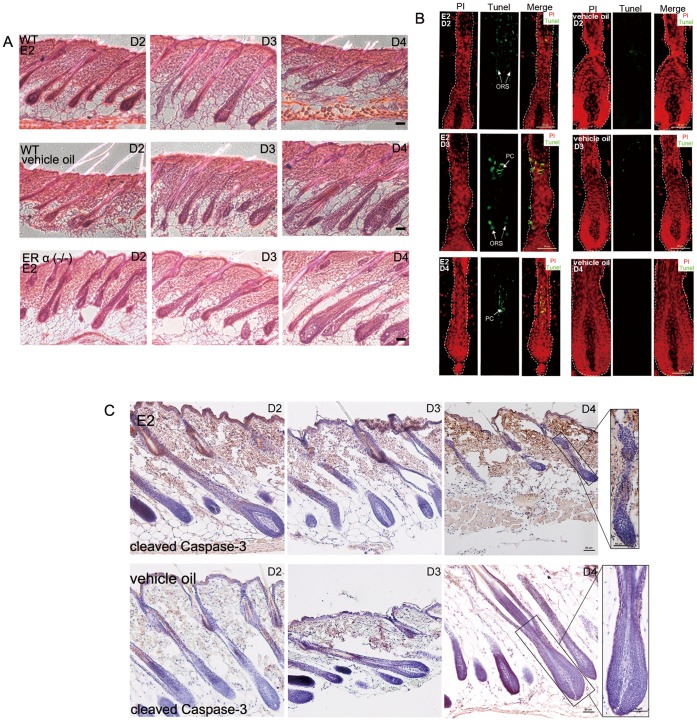
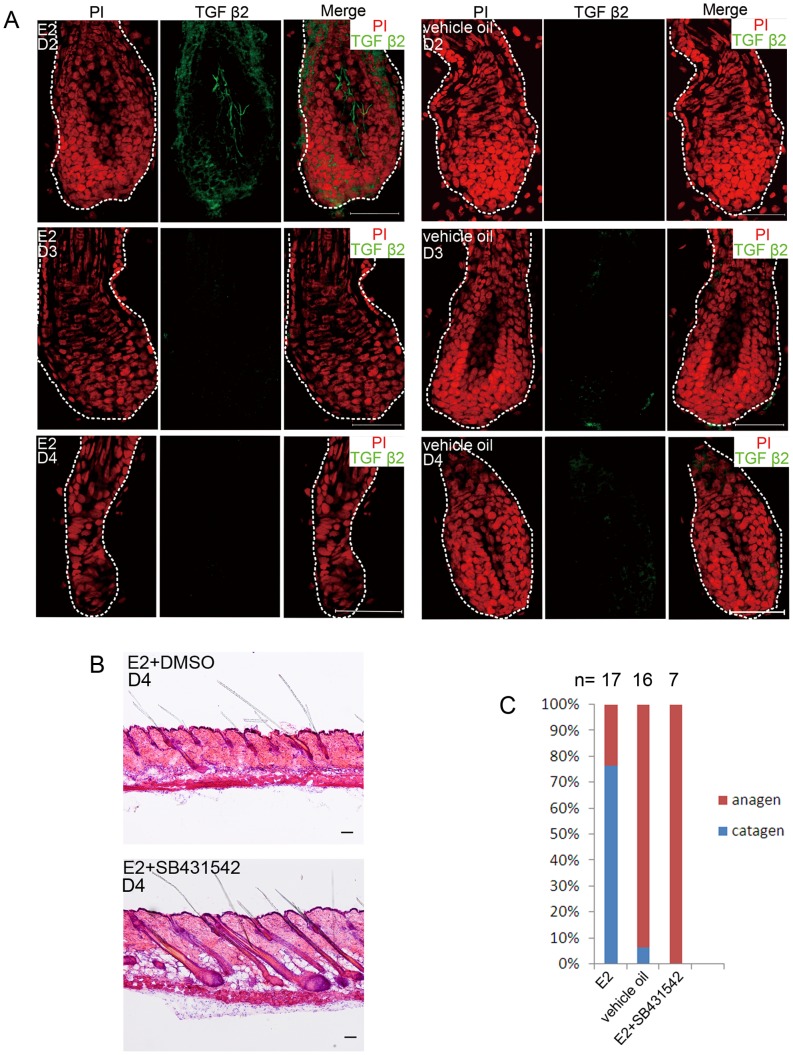
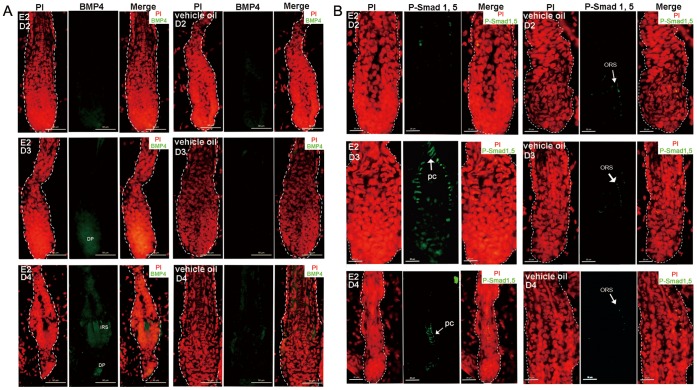
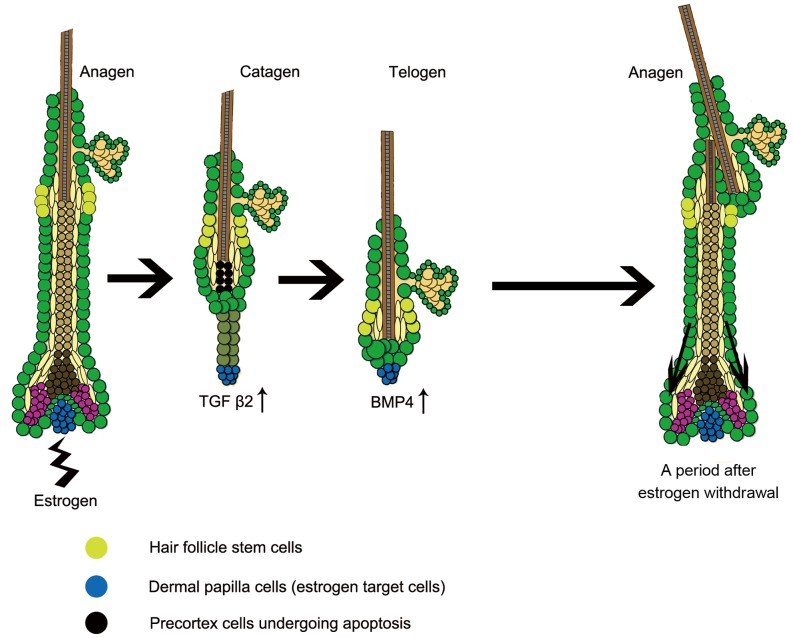
Estrogen dysregulation causes hair disorder. Clinical observations have demonstrated that estrogen raises the telogen/anagen ratio and inhibits hair shaft elongation of female scalp hair follicles. In spite of these clinical insights, the properties of estrogen on hair follicles are poorly dissected. In the present study, we show that estrogen induced apoptosis of precortex cells and caused premature catagen by up-regulation of TGF β2. Immediately after the premature catagen, the expression of anagen chalone BMP4 increased. The up-regulation of BMP4 may further function to prevent anagen transition and maintain telogen. Interestingly, the hair follicle stem cell niche was not destructed during these drastic structural changes caused by estrogen. Additionally, dermal papilla cells, the estrogen target cells in hair follicles, kept their signature gene expressions as well as their hair inductive potential after estrogen treatment. Retention of the characteristics of both hair follicle stem cells and dermal papilla cells determined the reversibility of the hair cycle suppression. These results indicated that estrogen causes reversible hair cycle retardation by inducing premature catagen and maintaining telogen.
Conflict of interest statement
Figures
Similar articles
-
Towards dissecting the pathogenesis of retinoid-induced hair loss: all-trans retinoic acid induces premature hair follicle regression (catagen) by upregulation of transforming growth factor-beta2 in the dermal papilla.J Invest Dermatol. 2005 Jun;124(6):1119-26. doi: 10.1111/j.0022-202X.2005.23686.x. J Invest Dermatol. 2005. PMID: 15955085
-
HNG, A Humanin Analogue, Promotes Hair Growth by Inhibiting Anagen-to-Catagen Transition.Int J Mol Sci. 2020 Jun 26;21(12):4553. doi: 10.3390/ijms21124553. Int J Mol Sci. 2020. PMID: 32604799 Free PMC article.
-
Interferon-gamma is a potent inducer of catagen-like changes in cultured human anagen hair follicles.Br J Dermatol. 2005 Apr;152(4):623-31. doi: 10.1111/j.1365-2133.2005.06453.x. Br J Dermatol. 2005. PMID: 15840090
-
[Regulation of hair follicle cycle].Morfologiia. 2014;146(5):83-7. Morfologiia. 2014. PMID: 25823297 Review. Russian.
-
[Research progress in regulation of hair growth by dermal adipose tissue].Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2024 May 15;38(5):626-632. doi: 10.7507/1002-1892.202402092. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2024. PMID: 38752252 Free PMC article. Review. Chinese.
Cited by
-
The role of CYP19A1 and ESR2 gene polymorphisms in female androgenetic alopecia in the Polish population.Postepy Dermatol Alergol. 2022 Aug;39(4):708-713. doi: 10.5114/ada.2021.108429. Epub 2021 Aug 16. Postepy Dermatol Alergol. 2022. PMID: 36090736 Free PMC article.
-
Postpartum Telogen Effluvium Unmasking Traction Alopecia.Skin Appendage Disord. 2022 Jul;8(4):328-332. doi: 10.1159/000521705. Epub 2022 Jan 28. Skin Appendage Disord. 2022. PMID: 35983466 Free PMC article.
-
Adipocytes in skin health and disease.Cold Spring Harb Perspect Med. 2014 Mar 1;4(3):a015271. doi: 10.1101/cshperspect.a015271. Cold Spring Harb Perspect Med. 2014. PMID: 24591537 Free PMC article. Review.
-
Epidermal development in mammals: key regulators, signals from beneath, and stem cells.Int J Mol Sci. 2013 May 24;14(6):10869-95. doi: 10.3390/ijms140610869. Int J Mol Sci. 2013. PMID: 23708093 Free PMC article. Review.
-
Estrogen regulates the expression of retinoic acid synthesis enzymes and binding proteins in mouse skin.Nutr Res. 2021 Oct;94:10-24. doi: 10.1016/j.nutres.2021.08.002. Epub 2021 Aug 24. Nutr Res. 2021. PMID: 34571215 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
