

Structural insights into the effector-immunity system Tse1/Tsi1 from Pseudomonas aeruginosa
- PMID: 22792331
- PMCID: PMC3391265
- DOI: 10.1371/journal.pone.0040453
Structural insights into the effector-immunity system Tse1/Tsi1 from Pseudomonas aeruginosa
Abstract
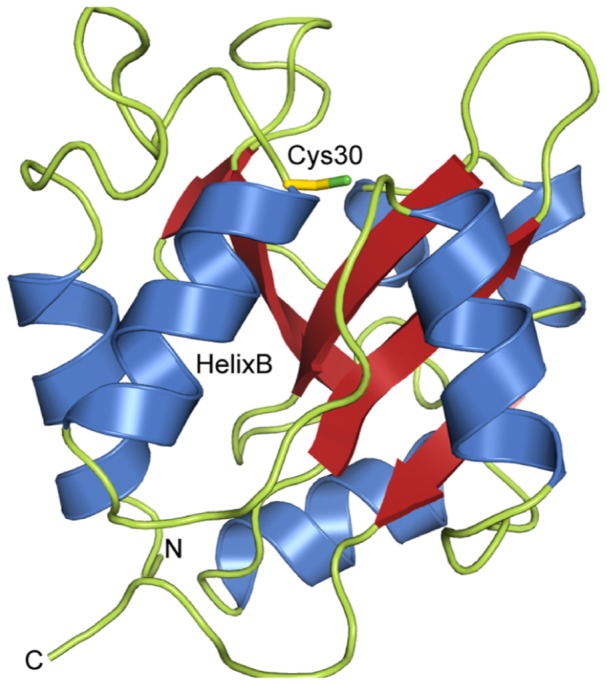
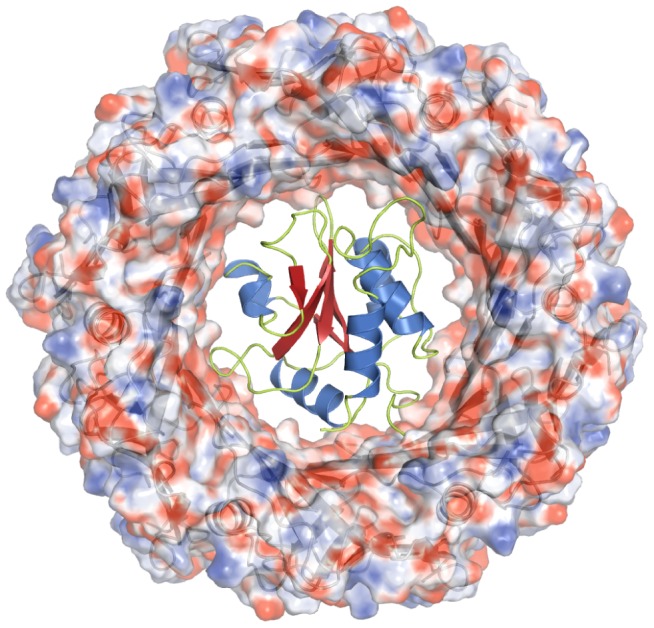
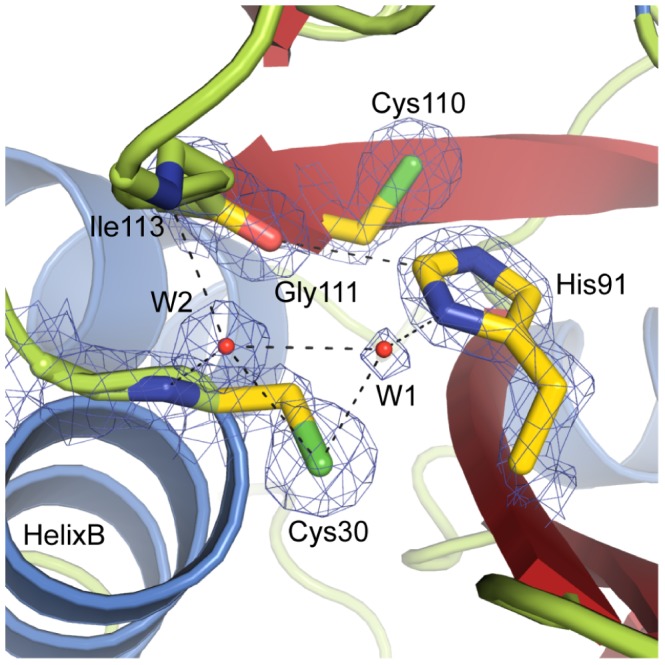
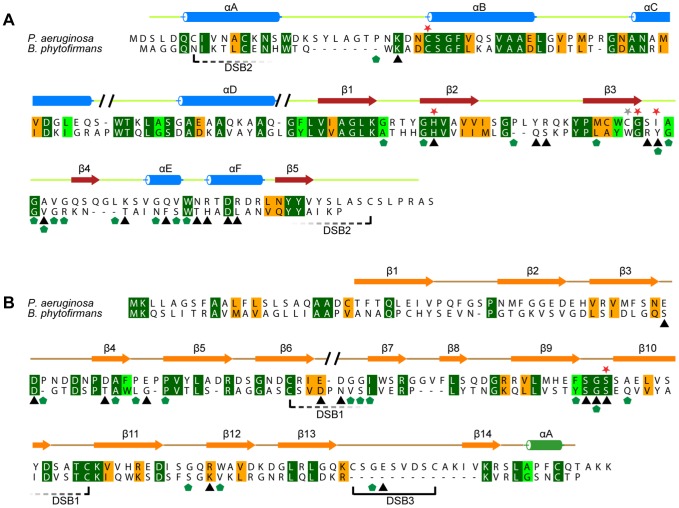
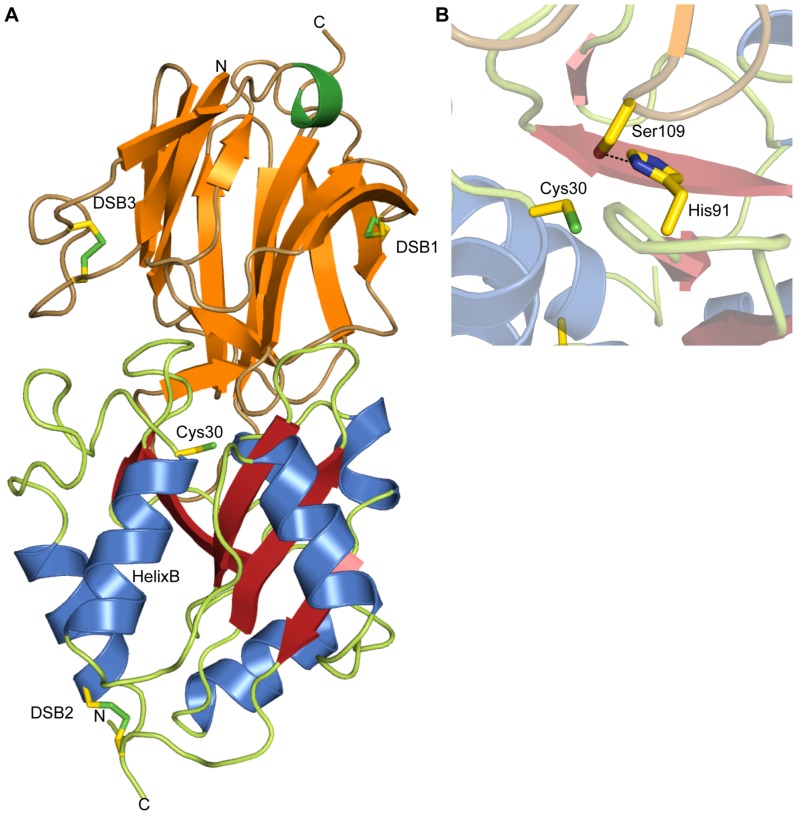
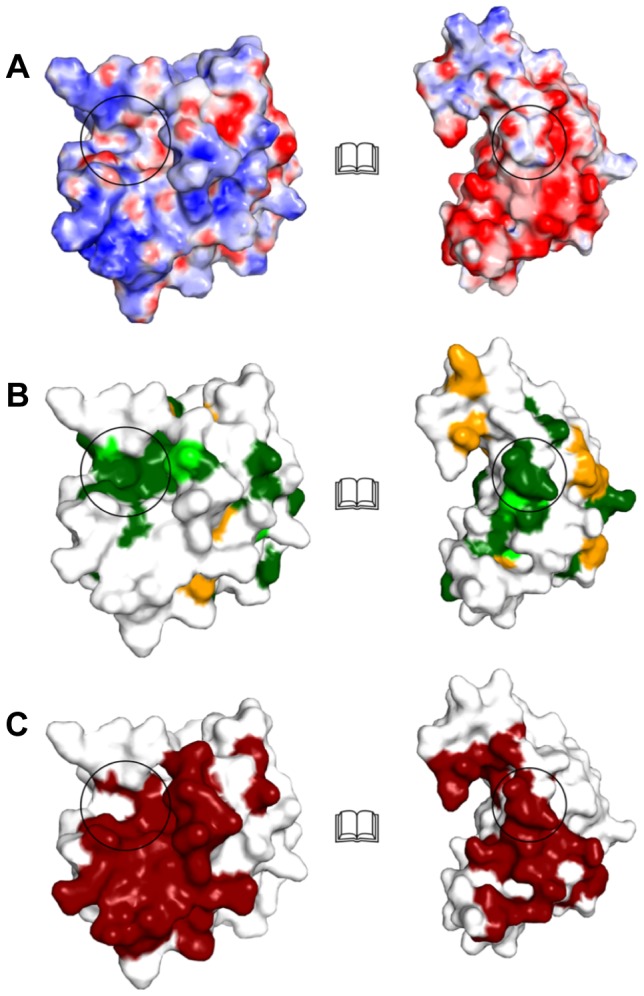
During an interbacterial battle, the type-6-secretion-system (T6SS) of the human pathogen Pseudomonas aeruginosa injects the peptidoglycan(PG)-hydrolase Tse1 into the periplasm of gram-negative enemy cells and induces their lysis. However, for its own benefit, P. aeruginosa produces and transports the immunity-protein Tsi1 into its own periplasm where in prevents accidental exo- and endogenous intoxication. Here we present the high-resolution X-ray crystal structure of the lytic enzyme Tse1 and describe the mechanism by which Tse1 cleaves the γ-D-glutamyl-l-meso-diaminopimelic acid amide bond of crosslinked PG. Tse1 belongs to the superfamily of N1pC/P60 peptidases but is unique among described members of this family of which the structure was described, since it is a single domain protein without any putative localization domain. Most importantly, we present the crystal structure of Tse1 bound to its immunity-protein Tsi1 as well and describe the mechanism of enzyme inhibition. Tsi1 occludes the active site of Tse1 and abolishes its enzyme activity by forming a hydrogen bond to a catalytically important histidine residue in Tse1. Based on our structural findings in combination with a bioinfomatic approach we also identified a related system in Burkholderia phytofirmans. Not only do our findings point to a common catalytic mechanism of the Tse1 PG-hydrolases, but we can also show that it is distinct from other members of this superfamily. Furthermore, we provide strong evidence that the mechanism of enzyme inhibition between Tsi1 orthologues is conserved. This work is the first structural description of an entire effector/immunity pair injected by the T6SS system. Moreover, it is also the first example of a member of the N1pC/P60 superfamily which becomes inhibited upon binding to its cognate immunity protein.
Conflict of interest statement
Figures
Similar articles
-
Complex structure of type VI peptidoglycan muramidase effector and a cognate immunity protein.Acta Crystallogr D Biol Crystallogr. 2013 Oct;69(Pt 10):1889-900. doi: 10.1107/S090744491301576X. Epub 2013 Sep 20. Acta Crystallogr D Biol Crystallogr. 2013. PMID: 24100309 Free PMC article.
-
Structural insight into how Pseudomonas aeruginosa peptidoglycanhydrolase Tse1 and its immunity protein Tsi1 function.Biochem J. 2012 Dec 1;448(2):201-11. doi: 10.1042/BJ20120668. Biochem J. 2012. PMID: 22931054
-
Structural insights into the Pseudomonas aeruginosa type VI virulence effector Tse1 bacteriolysis and self-protection mechanisms.J Biol Chem. 2012 Aug 3;287(32):26911-20. doi: 10.1074/jbc.M112.368043. Epub 2012 Jun 14. J Biol Chem. 2012. PMID: 22700987 Free PMC article.
-
Crystal structure of type VI effector Tse1 from Pseudomonas aeruginosa.FEBS Lett. 2012 Sep 21;586(19):3193-9. doi: 10.1016/j.febslet.2012.06.036. Epub 2012 Jun 29. FEBS Lett. 2012. PMID: 22750141
-
Structure of a peptidoglycan amidase effector targeted to Gram-negative bacteria by the type VI secretion system.Cell Rep. 2012 Jun 28;1(6):656-64. doi: 10.1016/j.celrep.2012.05.016. Epub 2012 May 31. Cell Rep. 2012. PMID: 22813741 Free PMC article.
Cited by
-
Type VI secretion system effectors: poisons with a purpose.Nat Rev Microbiol. 2014 Feb;12(2):137-48. doi: 10.1038/nrmicro3185. Epub 2014 Jan 2. Nat Rev Microbiol. 2014. PMID: 24384601 Free PMC article. Review.
-
Complex structure of type VI peptidoglycan muramidase effector and a cognate immunity protein.Acta Crystallogr D Biol Crystallogr. 2013 Oct;69(Pt 10):1889-900. doi: 10.1107/S090744491301576X. Epub 2013 Sep 20. Acta Crystallogr D Biol Crystallogr. 2013. PMID: 24100309 Free PMC article.
-
Structural Insights into the Effector - Immunity System Tae4/Tai4 from Salmonella typhimurium.PLoS One. 2013 Jun 27;8(6):e67362. doi: 10.1371/journal.pone.0067362. Print 2013. PLoS One. 2013. PMID: 23826277 Free PMC article.
-
A view to a kill: the bacterial type VI secretion system.Cell Host Microbe. 2014 Jan 15;15(1):9-21. doi: 10.1016/j.chom.2013.11.008. Epub 2013 Dec 11. Cell Host Microbe. 2014. PMID: 24332978 Free PMC article. Review.
-
The T6SSs of Pseudomonas aeruginosa Strain PAO1 and Their Effectors: Beyond Bacterial-Cell Targeting.Front Cell Infect Microbiol. 2016 Jun 9;6:61. doi: 10.3389/fcimb.2016.00061. eCollection 2016. Front Cell Infect Microbiol. 2016. PMID: 27376031 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
