

Intact memory for irrelevant information impairs perception in amnesia
- PMID: 22794269
- PMCID: PMC3657172
- DOI: 10.1016/j.neuron.2012.05.014
Intact memory for irrelevant information impairs perception in amnesia
Abstract
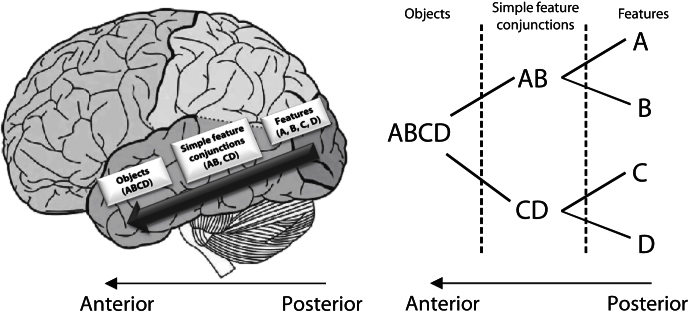
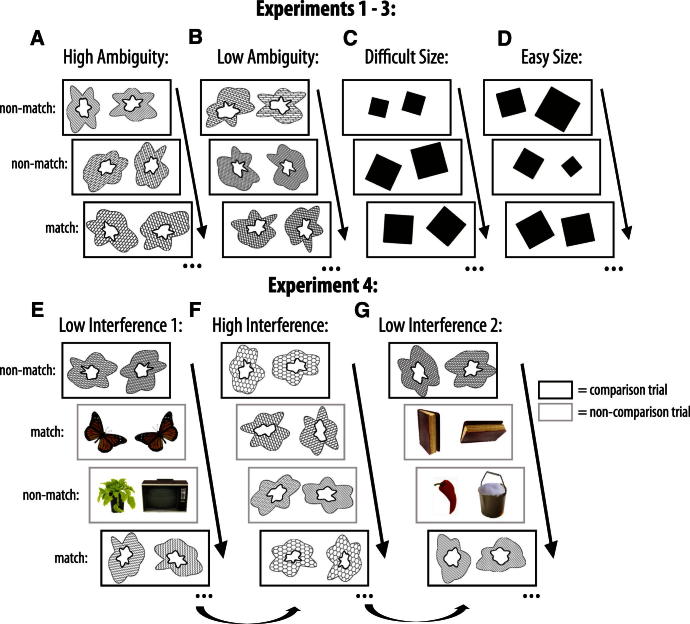
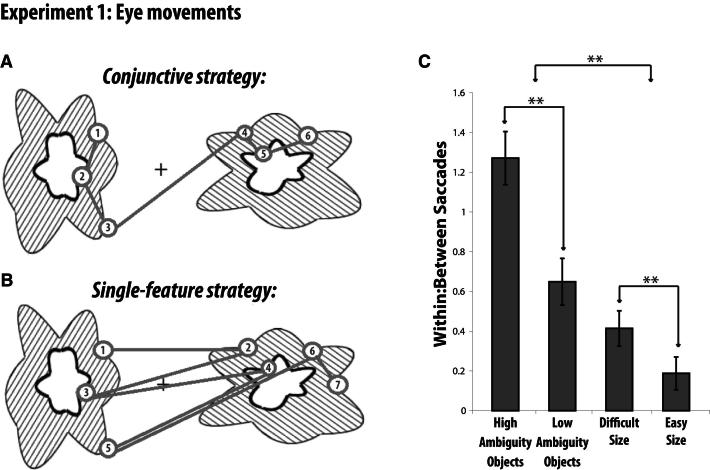
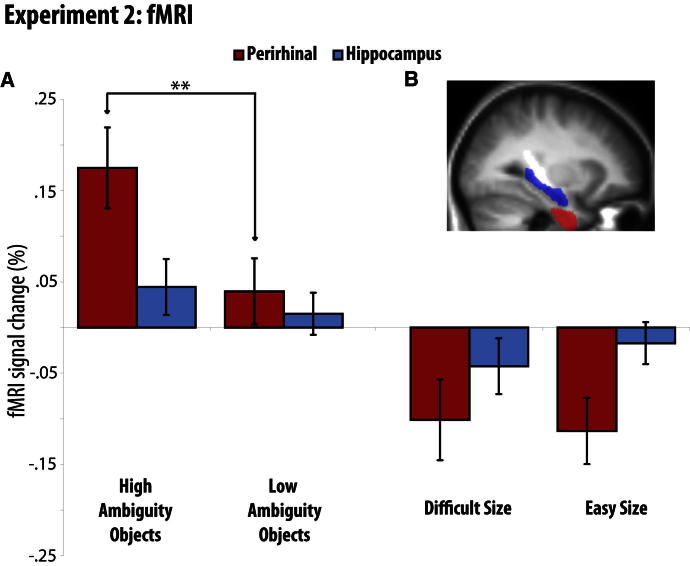
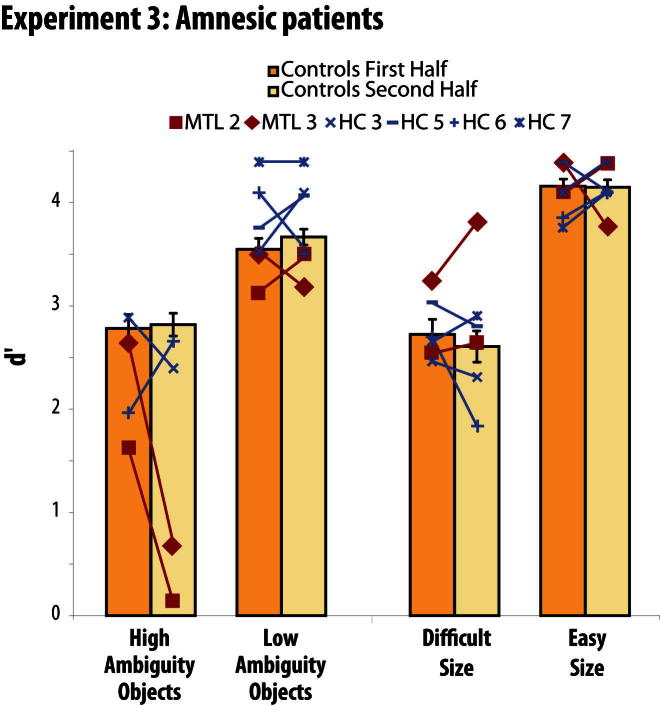
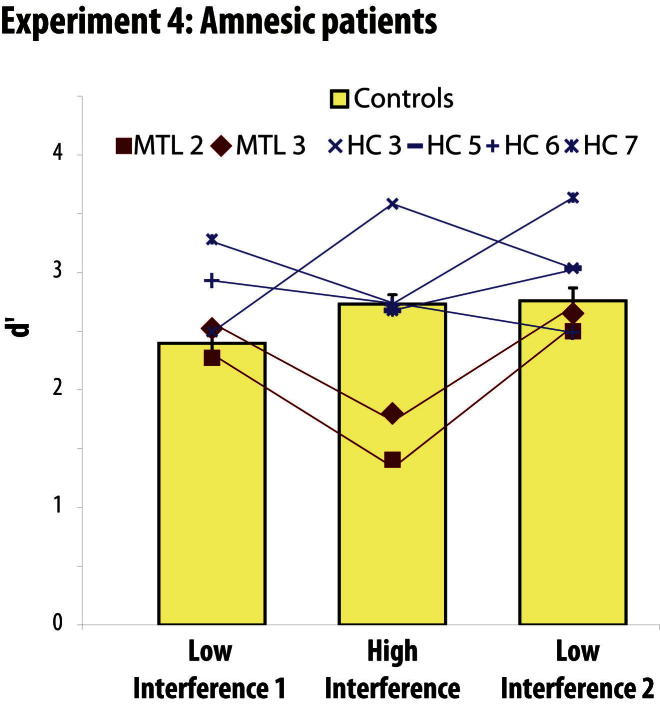
Memory and perception have long been considered separate cognitive processes, and amnesia resulting from medial temporal lobe (MTL) damage is thought to reflect damage to a dedicated memory system. Recent work has questioned these views, suggesting that amnesia can result from impoverished perceptual representations in the MTL, causing an increased susceptibility to interference. Using a perceptual matching task for which fMRI implicated a specific MTL structure, the perirhinal cortex, we show that amnesics with MTL damage including the perirhinal cortex, but not those with damage limited to the hippocampus, were vulnerable to object-based perceptual interference. Importantly, when we controlled such interference, their performance recovered to normal levels. These findings challenge prevailing conceptions of amnesia, suggesting that effects of damage to specific MTL regions are better understood not in terms of damage to a dedicated declarative memory system, but in terms of impoverished representations of the stimuli those regions maintain.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
Comment in
-
It's all coming back to me now: perception and memory in amnesia.Neuron. 2012 Jul 12;75(1):8-10. doi: 10.1016/j.neuron.2012.06.019. Neuron. 2012. PMID: 22794256
References
-
- Barense M.D., Gaffan D., Graham K.S. The human medial temporal lobe processes online representations of complex objects. Neuropsychologia. 2007;45:2963–2974. - PubMed
-
- Barense M.D., Rogers T.T., Bussey T.J., Saksida L.M., Graham K.S. Influence of conceptual knowledge on visual object discrimination: insights from semantic dementia and MTL amnesia. Cereb. Cortex. 2010;20:2568–2582. - PubMed
-
- Barense M.D., Henson R.N., Graham K.S. Perception and conception: temporal lobe activity during complex discriminations of familiar and novel faces and objects. J. Cogn. Neurosci. 2011;23:3052–3067. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
