

Independence of repressive histone marks and chromatin compaction during senescent heterochromatic layer formation
- PMID: 22795131
- PMCID: PMC3701408
- DOI: 10.1016/j.molcel.2012.06.010
Independence of repressive histone marks and chromatin compaction during senescent heterochromatic layer formation
Abstract
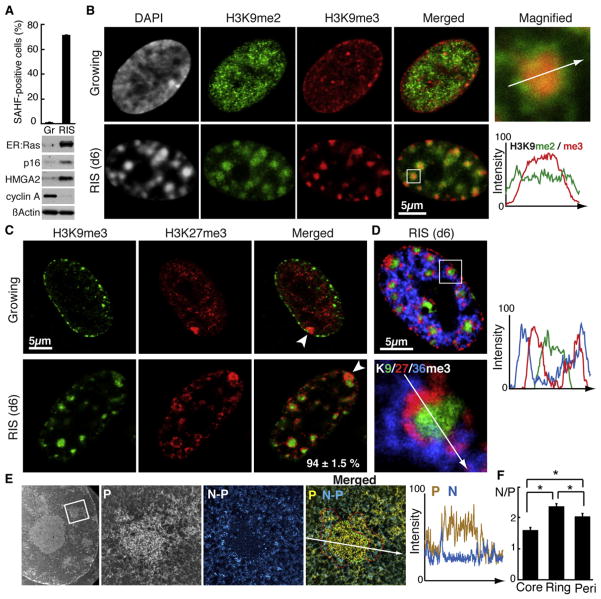
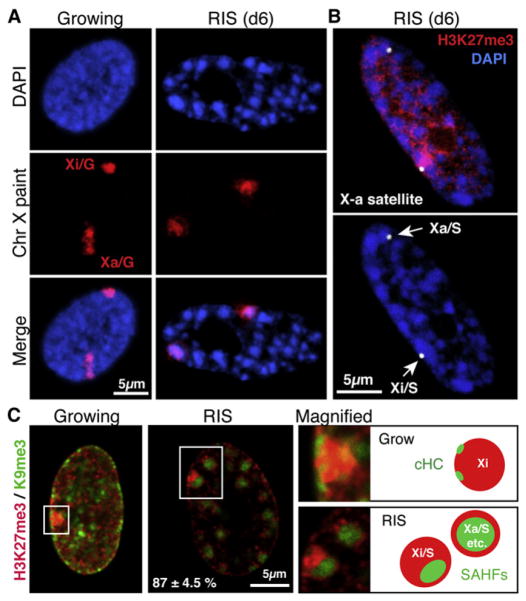
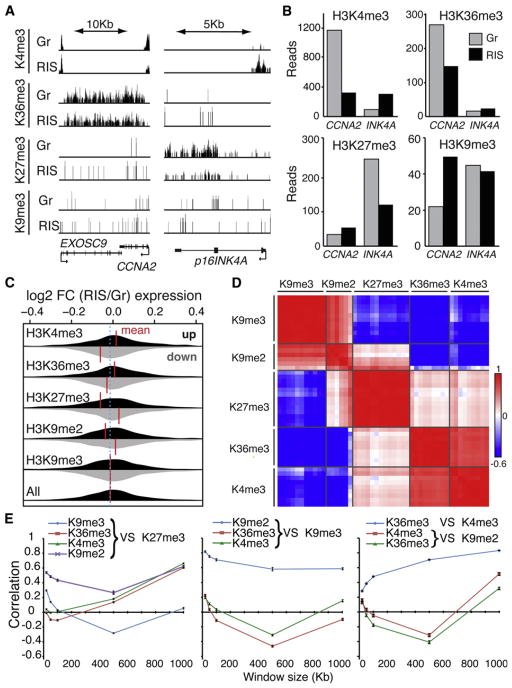
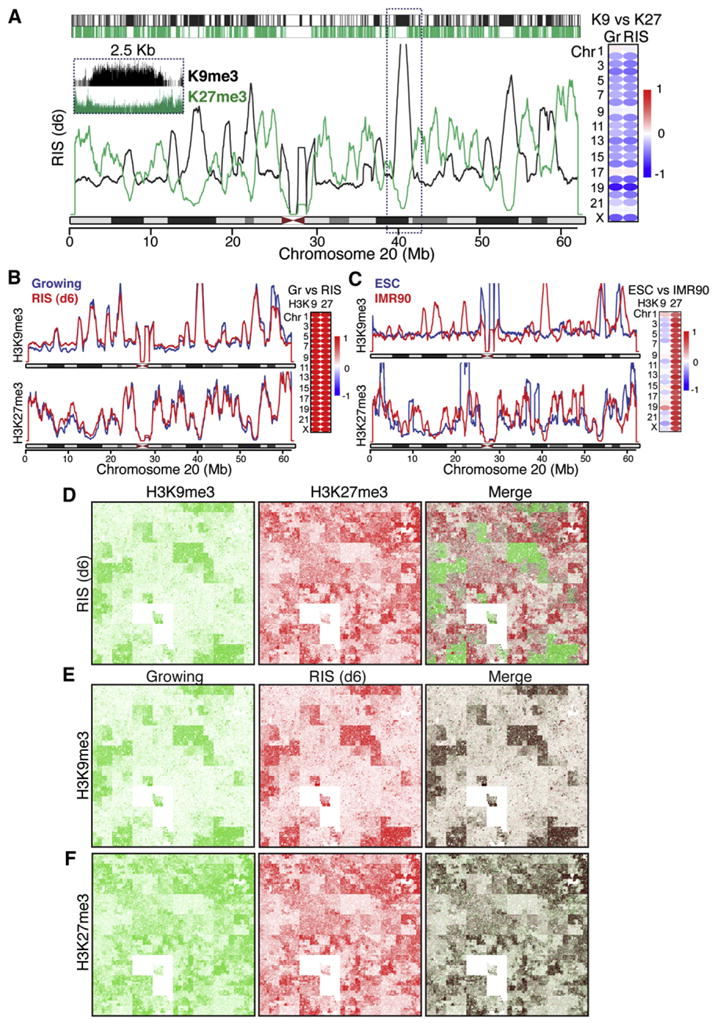
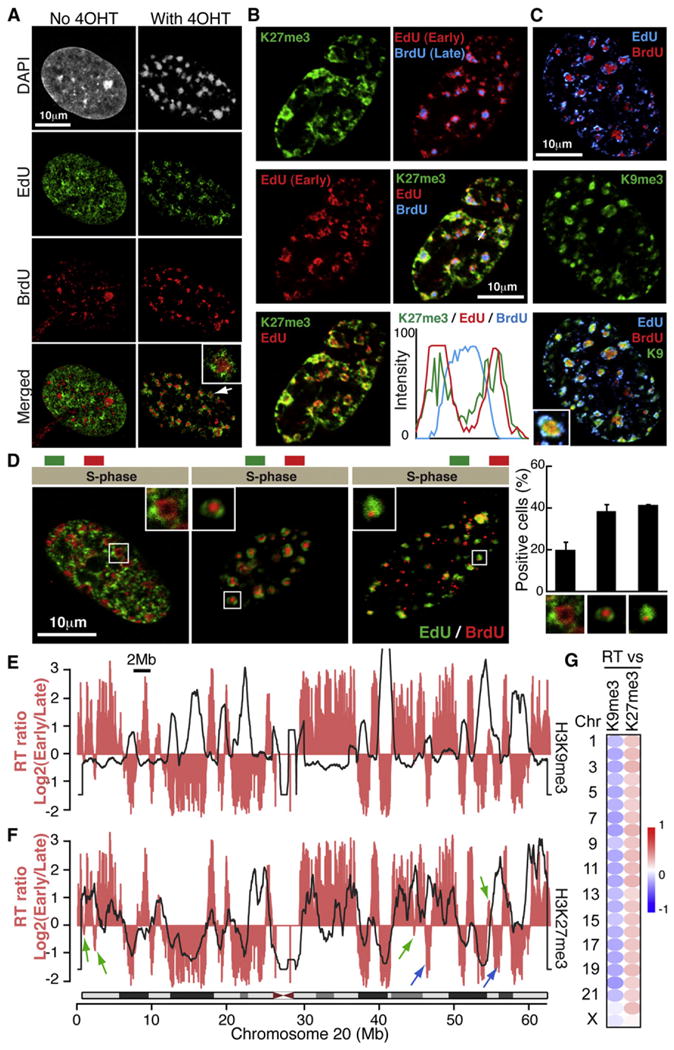
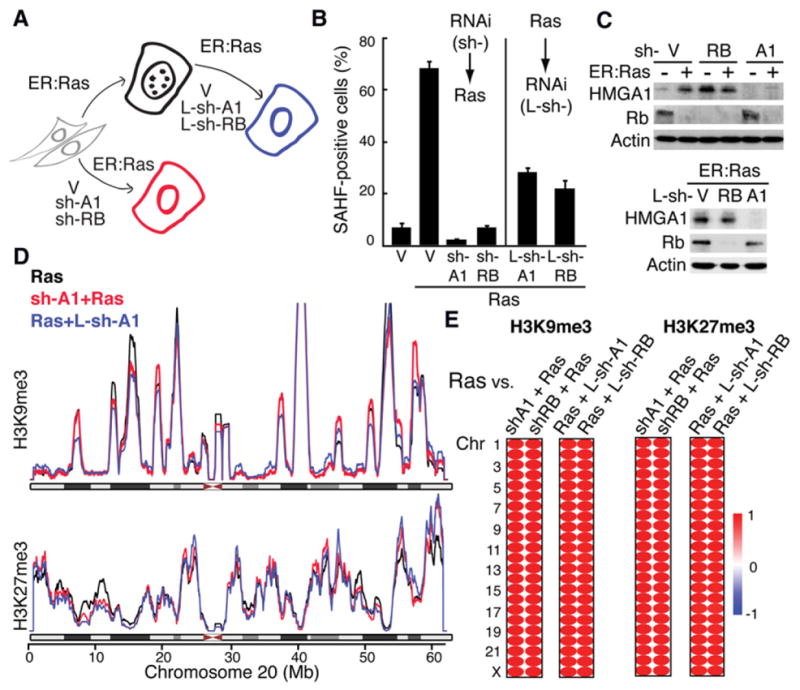
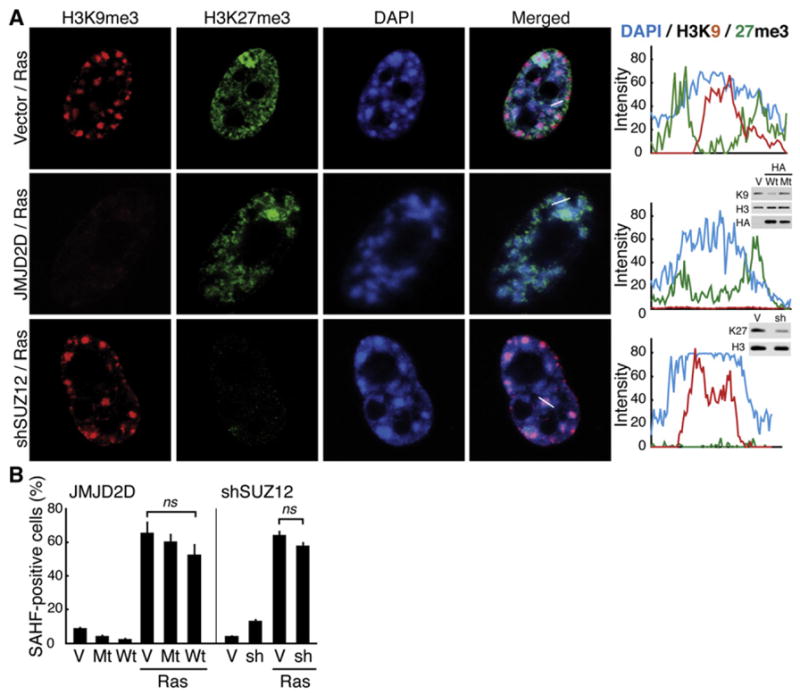
The expansion of repressive epigenetic marks has been implicated in heterochromatin formation during embryonic development, but the general applicability of this mechanism is unclear. Here we show that nuclear rearrangement of repressive histone marks H3K9me3 and H3K27me3 into nonoverlapping structural layers characterizes senescence-associated heterochromatic foci (SAHF) formation in human fibroblasts. However, the global landscape of these repressive marks remains unchanged upon SAHF formation, suggesting that in somatic cells, heterochromatin can be formed through the spatial repositioning of pre-existing repressively marked histones. This model is reinforced by the correlation of presenescent replication timing with both the subsequent layered structure of SAHFs and the global landscape of the repressive marks, allowing us to integrate microscopic and genomic information. Furthermore, modulation of SAHF structure does not affect the occupancy of these repressive marks, nor vice versa. These experiments reveal that high-order heterochromatin formation and epigenetic remodeling of the genome can be discrete events.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
- MC_U105185859/MRC_/Medical Research Council/United Kingdom
- P01 GM085354/GM/NIGMS NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- R01 AG016379/AG/NIA NIH HHS/United States
- R01 GM083337/GM/NIGMS NIH HHS/United States
- 202218/ERC_/European Research Council/International
- CAPMC/ CIHR/Canada
- GM085354/GM/NIGMS NIH HHS/United States
- 15603/CRUK_/Cancer Research UK/United Kingdom
- GM083337/GM/NIGMS NIH HHS/United States
- 15890/CRUK_/Cancer Research UK/United Kingdom
- A15603/CRUK_/Cancer Research UK/United Kingdom
- 14545/CRUK_/Cancer Research UK/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
