

Molecular force transduction by ion channels: diversity and unifying principles
- PMID: 22797911
- PMCID: PMC3434843
- DOI: 10.1242/jcs.092353
Molecular force transduction by ion channels: diversity and unifying principles
Abstract
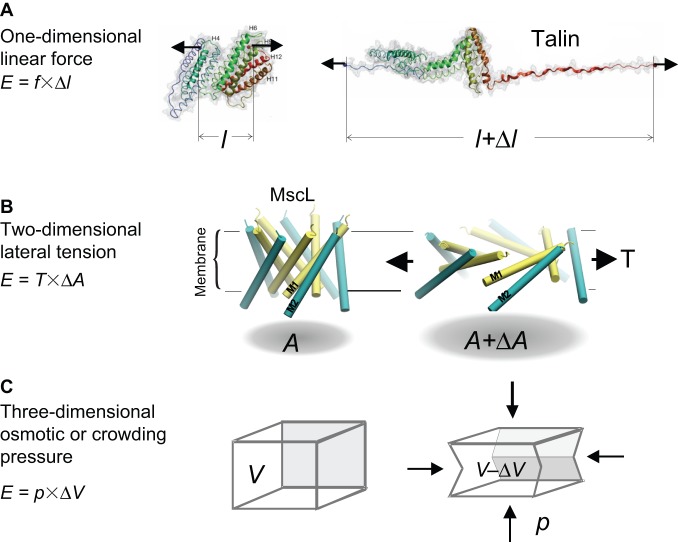
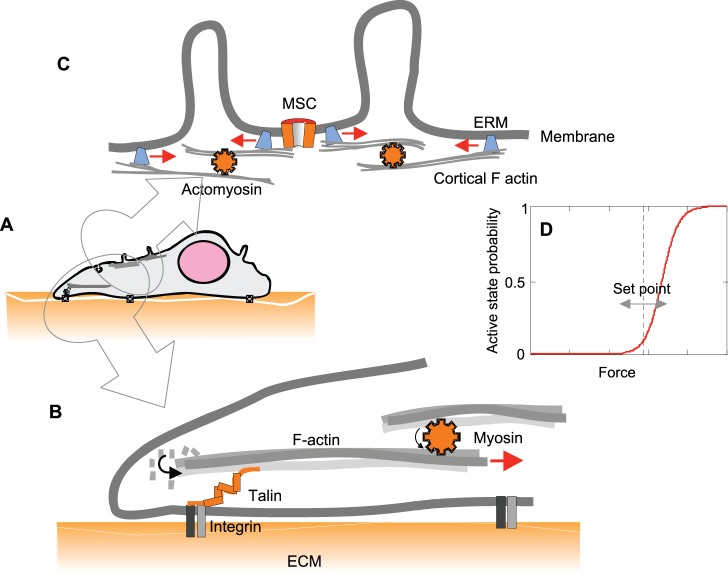
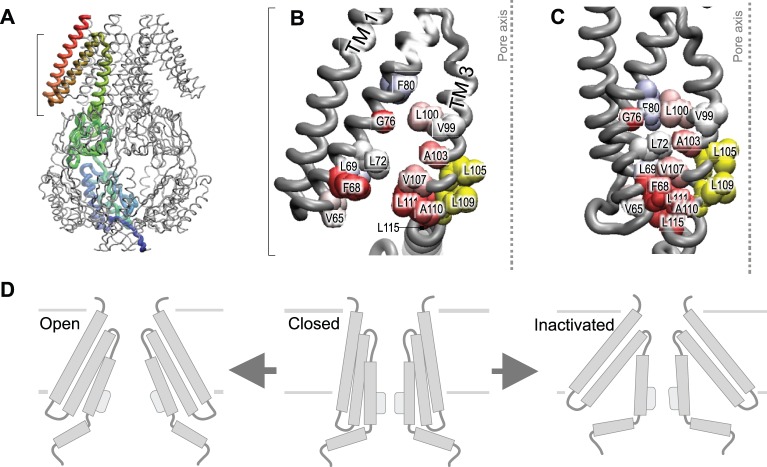
Cells perceive force through a variety of molecular sensors, of which the mechanosensitive ion channels are the most efficient and act the fastest. These channels apparently evolved to prevent osmotic lysis of the cell as a result of metabolite accumulation and/or external changes in osmolarity. From this simple beginning, nature developed specific mechanosensitive enzymes that allow us to hear, maintain balance, feel touch and regulate many systemic variables, such as blood pressure. For a channel to be mechanosensitive it needs to respond to mechanical stresses by changing its shape between the closed and open states. In that way, forces within the lipid bilayer or within a protein link can do work on the channel and stabilize its state. Ion channels have the highest turnover rates of all enzymes, and they can act as both sensors and effectors, providing the necessary fluxes to relieve osmotic pressure, shift the membrane potential or initiate chemical signaling. In this Commentary, we focus on the common mechanisms by which mechanical forces and the local environment can regulate membrane protein structure, and more specifically, mechanosensitive ion channels.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
