

S-glutathionylation of the Na,K-ATPase catalytic α subunit is a determinant of the enzyme redox sensitivity
- PMID: 22798075
- PMCID: PMC3442550
- DOI: 10.1074/jbc.M112.391094
S-glutathionylation of the Na,K-ATPase catalytic α subunit is a determinant of the enzyme redox sensitivity
Abstract
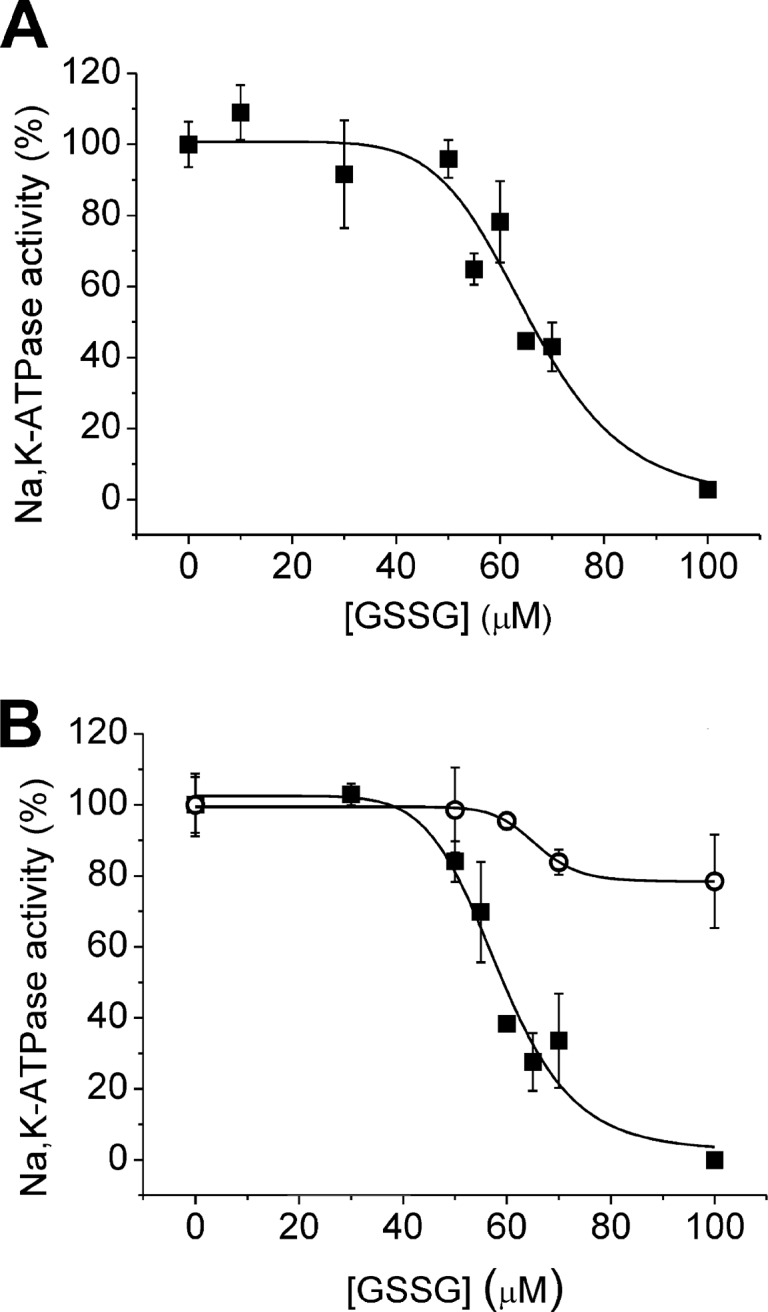
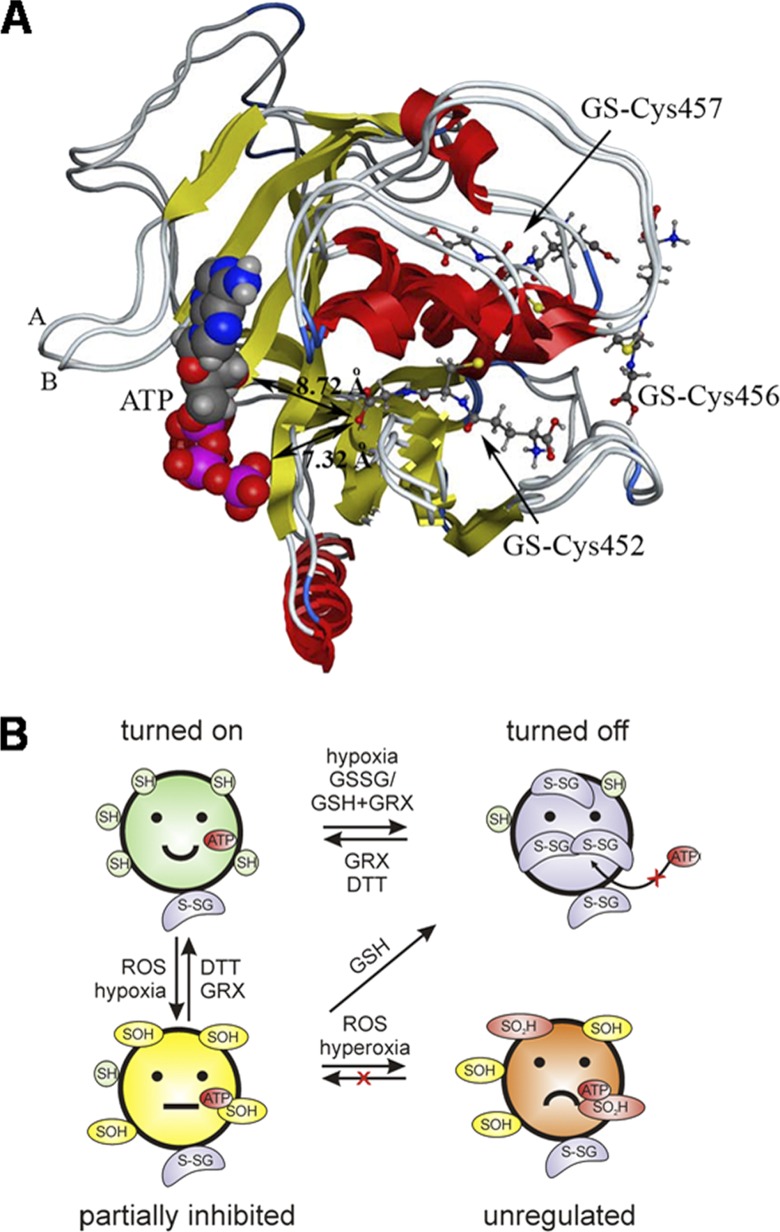
Na,K-ATPase is highly sensitive to changes in the redox state, and yet the mechanisms of its redox sensitivity remain unclear. We have explored the possible involvement of S-glutathionylation of the catalytic α subunit in redox-induced responses. For the first time, the presence of S-glutathionylated cysteine residues was shown in the α subunit in duck salt glands, rabbit kidneys, and rat myocardium. Exposure of the Na,K-ATPase to oxidized glutathione (GSSG) resulted in an increase in the number of S-glutathionylated cysteine residues. Increase in S-glutathionylation was associated with dose- and time-dependent suppression of the enzyme function up to its complete inhibition. The enzyme inhibition concurred with S-glutathionylation of the Cys-454, -458, -459, and -244. Upon binding of glutathione to these cysteines, the enzyme was unable to interact with adenine nucleotides. Inhibition of the Na,K-ATPase by GSSG did not occur in the presence of ATP at concentrations above 0.5 mm. Deglutathionylation of the α subunit catalyzed by glutaredoxin or dithiothreitol resulted in restoration of the Na,K-ATPase activity. Oxidation of regulatory cysteines made them inaccessible for glutathionylation but had no profound effect on the enzyme activity. Regulatory S-glutathionylation of the α subunit was induced in rat myocardium in response to hypoxia and was associated with oxidative stress and ATP depletion. S-Glutathionylation was followed by suppression of the Na,K-ATPase activity. The rat α2 isoform was more sensitive to GSSG than the α1 isoform. Our findings imply that regulatory S-glutathionylation of the catalytic subunit plays a key role in the redox-induced regulation of Na,K-ATPase activity.
Figures





References
-
- Blanco G., Mercer R. W. (1998) Isozymes of the Na-K-ATPase. Heterogeneity in structure, diversity in function. Am. J. Physiol. 275, F633–F650 - PubMed
-
- Glitsch H. G. (2001) Electrophysiology of the sodium-potassium-ATPase in cardiac cells. Physiol. Rev. 81, 1791–1826 - PubMed
-
- Geering K. (2008) Functional roles of Na,K-ATPase subunits. Curr. Opin. Nephrol. Hypertens. 17, 526–532 - PubMed
-
- Kaplan J. H. (2002) Biochemistry of Na,K-ATPase. Annu. Rev. Biochem. 71, 511–535 - PubMed
-
- Bogdanova A., Petrushanko I., Boldyrev A., Gassmann M. (2006) Oxygen- and redox-induced regulation of the Na/K-ATPase. Curr. Enzyme Inhibition 2, 37–59
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
