

The potassium channel, Kir3.4 participates in angiotensin II-stimulated aldosterone production by a human adrenocortical cell line
- PMID: 22798349
- PMCID: PMC3423613
- DOI: 10.1210/en.2012-1241
The potassium channel, Kir3.4 participates in angiotensin II-stimulated aldosterone production by a human adrenocortical cell line
Abstract
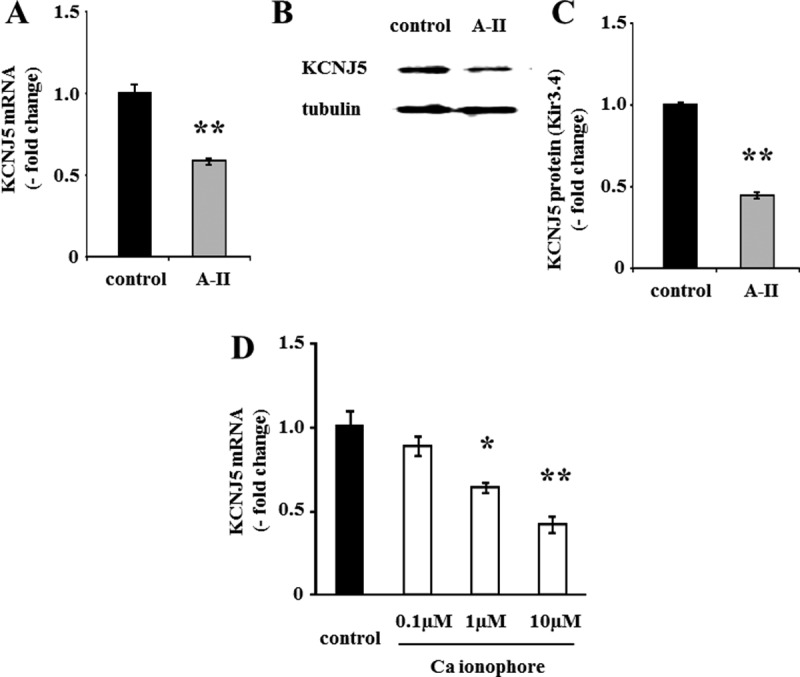
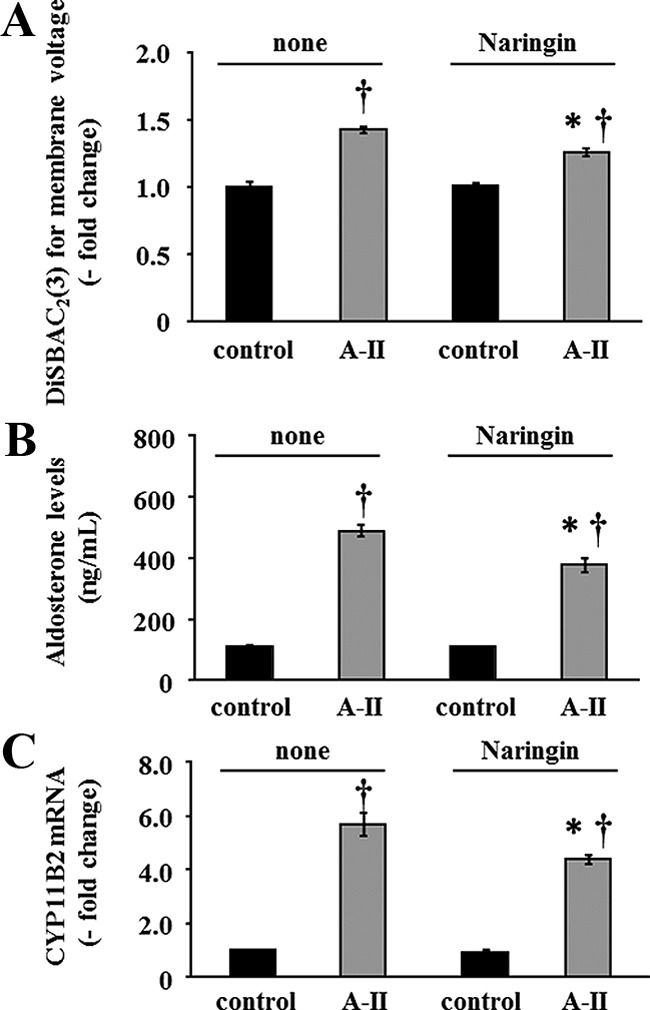
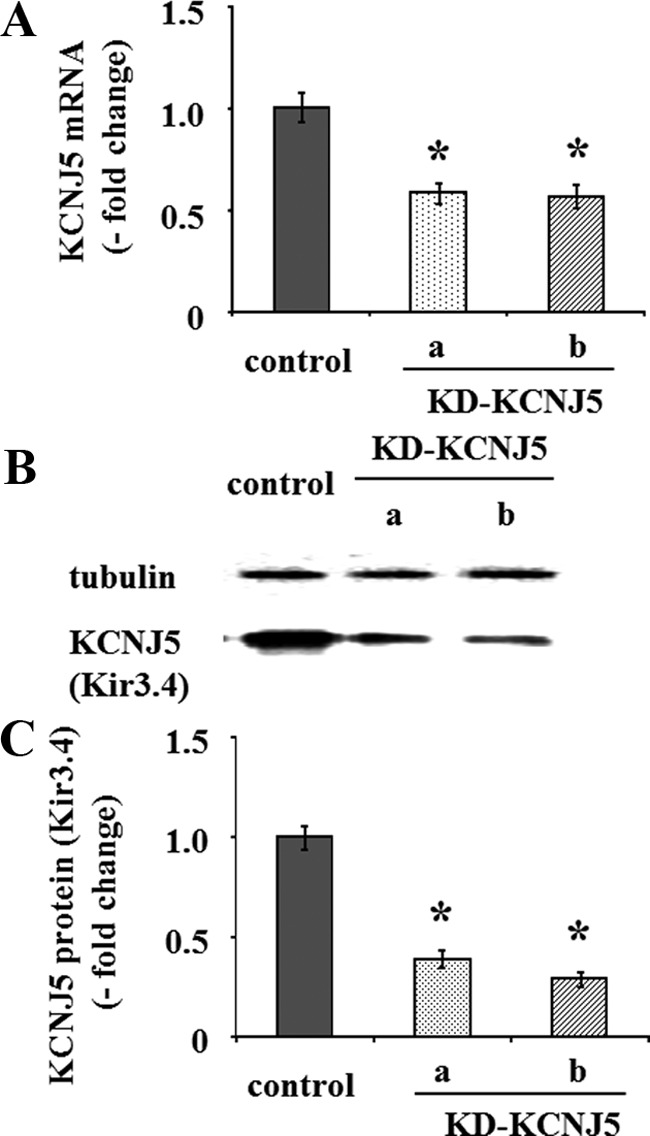
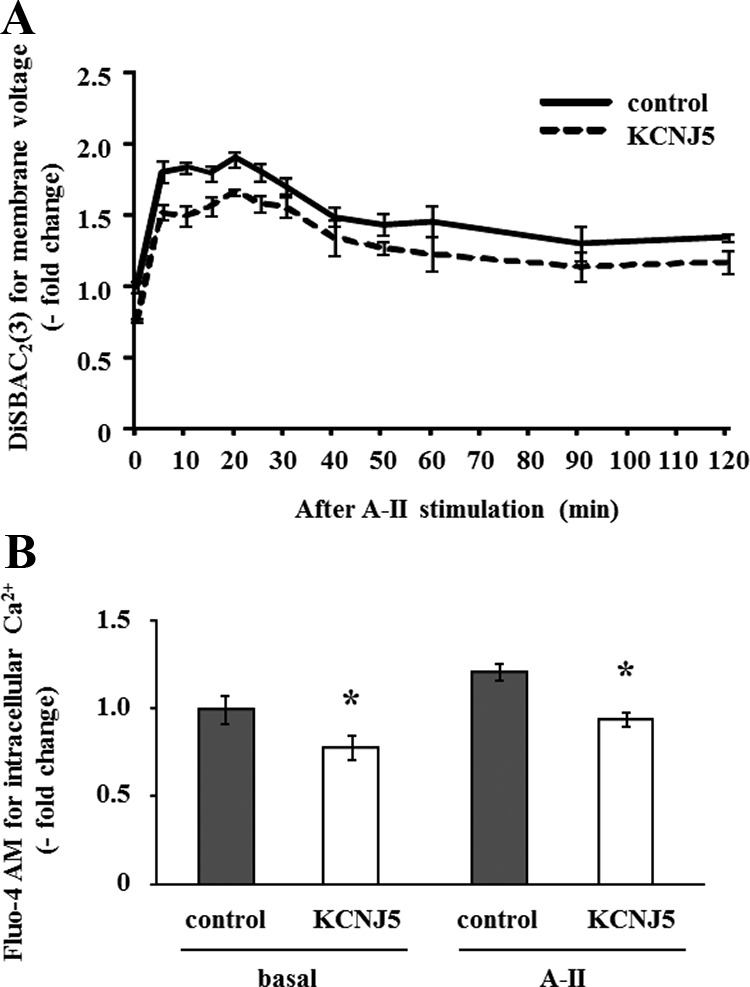
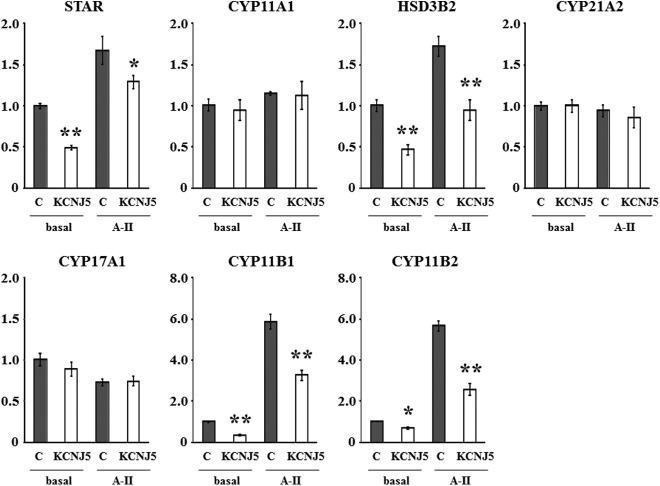
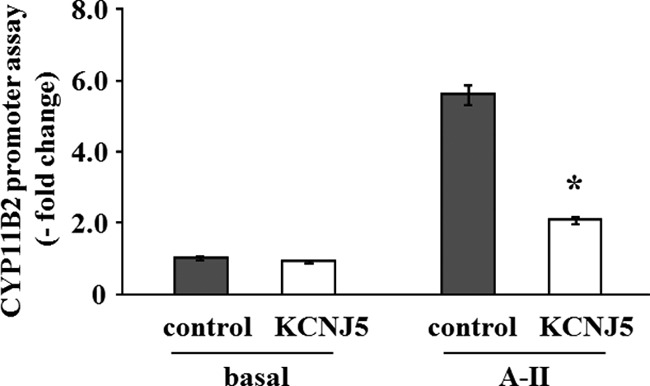
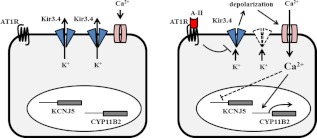
Angiotensin II (A-II) regulation of aldosterone secretion is initiated by inducing cell membrane depolarization, thereby increasing intracellular calcium and activating the calcium calmodulin/calmodulin kinase cascade. Mutations in the selectivity filter of the KCNJ5 gene coding for inward rectifying potassium channel (Kir)3.4 has been found in about one third of aldosterone-producing adenomas. These mutations result in loss of selectivity of the inward rectifying current for potassium, which causes membrane depolarization and opening of calcium channels and activation of the calcium calmodulin/calmodulin kinase cascade and results in an increase in aldosterone secretion. In this study we show that A-II and a calcium ionophore down-regulate the expression of KCNJ5 mRNA and protein. Activation of Kir3.4 by naringin inhibits A-II-stimulated membrane voltage and aldosterone secretion. Overexpression of KCNJ5 in the HAC15 cells using a lentivirus resulted in a decrease in membrane voltage, intracellular calcium, expression of steroidogenic acute regulatory protein, 3-β-hydroxysteroid dehydrogenase 3B2, cytochrome P450 11B1 and cytochrome P450 11B2 mRNA, and aldosterone synthesis. In conclusion, A-II appears to stimulate aldosterone secretion by depolarizing the membrane acting in part through the regulation of the expression and activity of Kir3.4.
Figures

References
-
- Funder JW, Carey RM, Fardella C, Gomez-Sanchez CE, Mantero F, Stowasser M, Young WF, Jr, Montori VM. 2008. Case detection, diagnosis, and treatment of patients with primary aldosteronism: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab 93:3266–3281 - PubMed
-
- Milliez P, Girerd X, Plouin PF, Blacher J, Safar ME, Mourad JJ. 2005. Evidence for an increased rate of cardiovascular events in patients with primary aldosteronism. J Am Coll Cardiol 45:1243–1248 - PubMed
-
- Condon JC, Pezzi V, Drummond BM, Yin S, Rainey WE. 2002. Calmodulin-dependent kinase I regulates adrenal cell expression of aldosterone synthase. Endocrinology 143:3651–3657 - PubMed
-
- Otis M, Gallo-Payet N. 2006. Differential involvement of cytoskeleton and rho-guanosine 5′-triphosphatases in growth-promoting effects of angiotensin II in rat adrenal glomerulosa cells. Endocrinology 147:5460–5469 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
