

The representation of sound localization cues in the barn owl's inferior colliculus
- PMID: 22798945
- PMCID: PMC3394089
- DOI: 10.3389/fncir.2012.00045
The representation of sound localization cues in the barn owl's inferior colliculus
Abstract
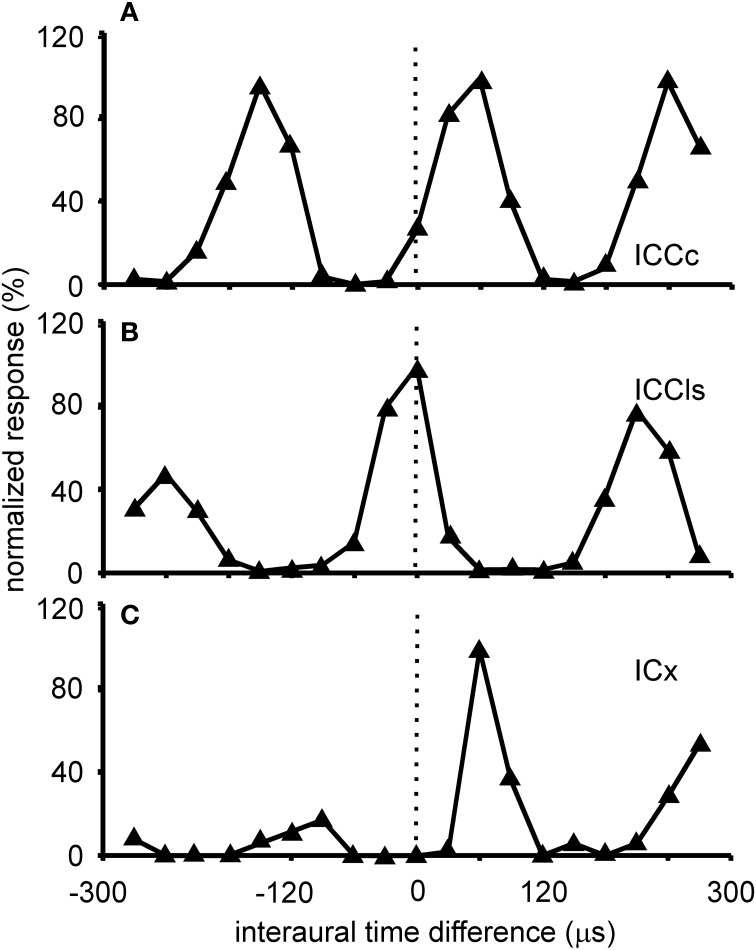
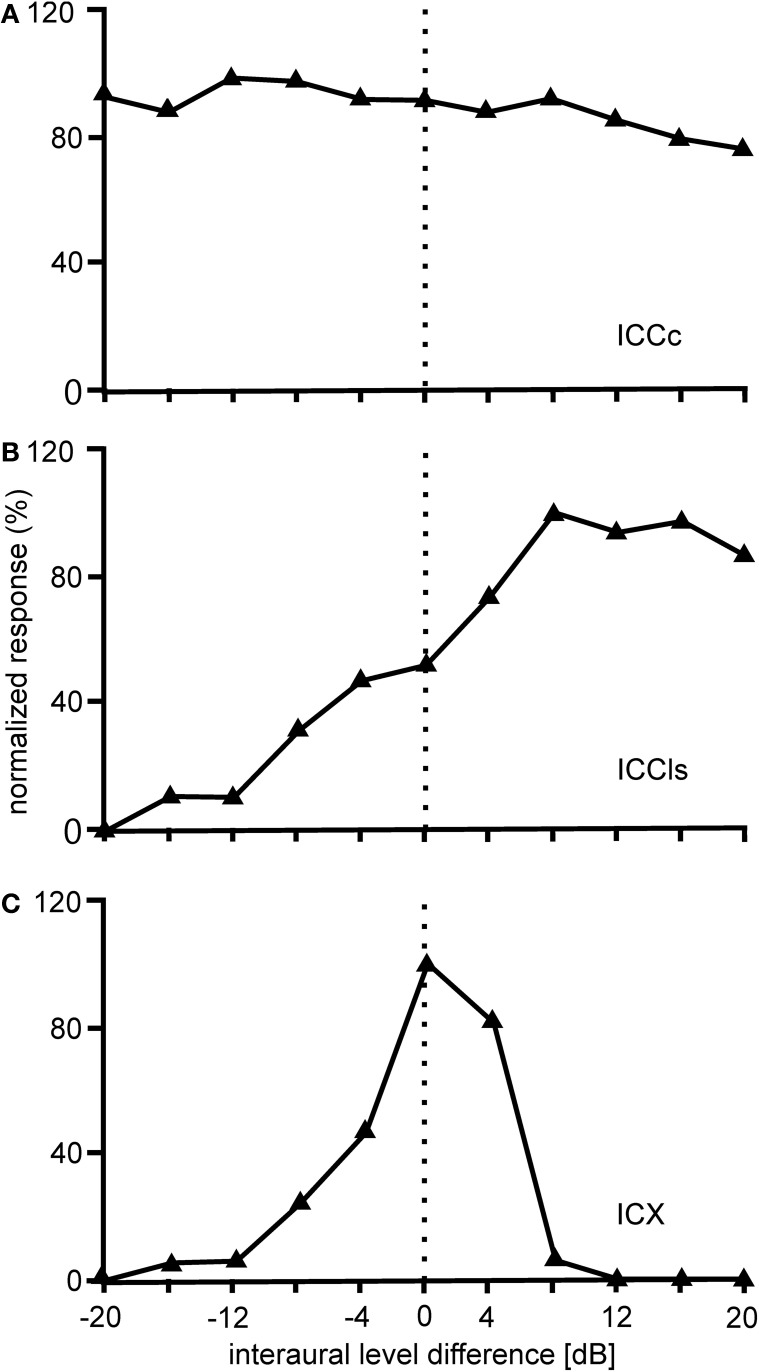
The barn owl is a well-known model system for studying auditory processing and sound localization. This article reviews the morphological and functional organization, as well as the role of the underlying microcircuits, of the barn owl's inferior colliculus (IC). We focus on the processing of frequency and interaural time (ITD) and level differences (ILD). We first summarize the morphology of the sub-nuclei belonging to the IC and their differentiation by antero- and retrograde labeling and by staining with various antibodies. We then focus on the response properties of neurons in the three major sub-nuclei of IC [core of the central nucleus of the IC (ICCc), lateral shell of the central nucleus of the IC (ICCls), and the external nucleus of the IC (ICX)]. ICCc projects to ICCls, which in turn sends its information to ICX. The responses of neurons in ICCc are sensitive to changes in ITD but not to changes in ILD. The distribution of ITD sensitivity with frequency in ICCc can only partly be explained by optimal coding. We continue with the tuning properties of ICCls neurons, the first station in the midbrain where the ITD and ILD pathways merge after they have split at the level of the cochlear nucleus. The ICCc and ICCls share similar ITD and frequency tuning. By contrast, ICCls shows sigmoidal ILD tuning which is absent in ICCc. Both ICCc and ICCls project to the forebrain, and ICCls also projects to ICX, where space-specific neurons are found. Space-specific neurons exhibit side peak suppression in ITD tuning, bell-shaped ILD tuning, and are broadly tuned to frequency. These neurons respond only to restricted positions of auditory space and form a map of two-dimensional auditory space. Finally, we briefly review major IC features, including multiplication-like computations, correlates of echo suppression, plasticity, and adaptation.
Keywords: adaptation; auditory; central nucleus of the inferior colliculus; frequency tuning; interaural level difference; interaural time difference; plasticity; sound localization.
Figures







References
-
- Aitkin L. M., Pettigrew J. D., Calford M. B., Philips S. C., Wise L. Z. (1985). Representation of stimulus azimuth by low-frequency neurons in inferior colliculus of the cat. J. Neurophysiol. 53, 43–59 - PubMed
LinkOut - more resources
Full Text Sources